Animal navigation is similar to conventional (formalized) navigation in at least three basic ways. First, it relies heavily on dead reckoning, the continual updating of position by summing successive small displacements (changes in position). In the limit (as the differences between successive positions are made arbitrarily small), this process is equivalent to obtaining the position vector by integrating the velocity vector with respect to time, which is why the process is also called path integration (see Gallistel 1990, chap. 4, for review of literature).
Second, it only occasionally takes a fix, that is, establishes position and heading (orientation) on a map using perceived distances and directions from mapped features of the terrain. While it is necessary from time to time to correct the inevitable cumulative error in dead reckoning, animals, like human navigators, often invert the process in the intervals between fixes, using their reckoned position and heading on their map to estimate their relation to surrounding features. In doing so, they appear to ignore the current testimony of their senses. Thus, when the dimensions of a familiar maze are altered, a rat moving rapidly through it collides with walls that come too soon, and turns into a wall where it expects an opening (Carr and Watson 1908). Indeed, it runs right over the pile of food that is its goal when it encounters that pile much sooner than expected (Stoltz and Lott 1964). Bats threading their way through obstacle courses bring their wings together over their head to squeeze through a gap that is no longer so narrow as to require this maneuver (Neuweiler and Möhres 1967).
And third, it places relatively minor reliance on beacon navigation, the following of sensory cues from a goal or its immediate surroundings. Animals of widely diverse species locate a goal by the goal's position relative to the general framework provided by the mapped terrain (see Gallistel 1990, chap. 5, for review), not by the sensory characteristics of the goal or its immediate surroundings. Indeed, when the two are placed in conflict, the animal goes to the place having the correct position in the larger framework, not to the place having the correct sensory characteristics. For example, if a chimpanzee sees food hidden in one of two differently colored containers whose positions are then surreptitiously interchanged, the chimpanzee searches for the food in the container at the correct location rather than in the one with the correct color (Tinkelpaugh 1932). In human toddlers, position also takes precedence over the characteristics of the container. When a toddler is misoriented within a rectangular room, it ignores the container in which it saw a toy hidden, but which it now mistakenly takes to be in the wrong location. It looks instead in an altogether different container, which it takes to be in the correct location, even though the child demonstrably remembers the appearance of the container in which it saw the toy hidden (Hermer and Spelke 1996).
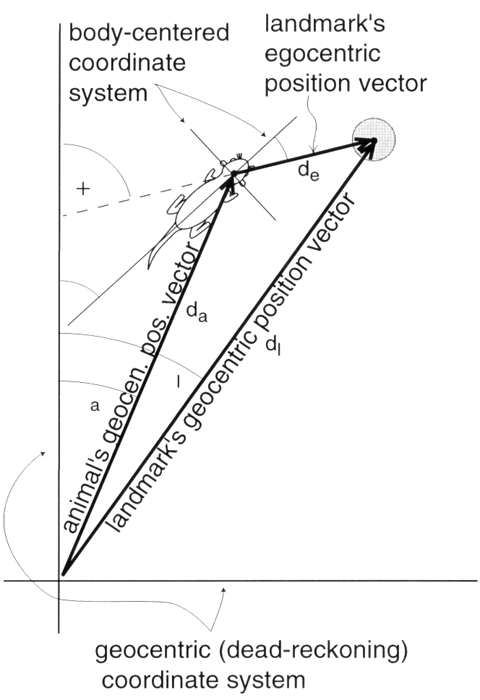
Figure 1. Conversion of egocentric position coordinates to a common geocentric framework. The egocentric position vector of the landmark, with angle β (bearing of landmark) and magnitude d1 is first rotated so that it has angle η + β (heading plus bearing), then added to the geocentric position vector of the animal (with compass angle γa and magnitude da), producing the geocentric position vector for the landmark (with compass angle γ1 and magnitude d1). (Slightly modified from figure 1 in Gallistel and Cramer 1996. Used by permission of the authors and publisher.)
The sensory cues from the goal itself appear to control approach behavior only in the final approach to the goal, and even then, only when the goal is in approximately the correct location. The same is true for the nearby landmarks with respect to which an animal more precisely locates its goal. It uses them as aids to locating its goal only if they occupy approximately the correct place in the larger framework. Bees readily learn to search for food on one side of a landmark in one location but on the opposite side of the identical landmark placed in a different location (Collett and Kelber 1988). In effect, location confers identity, rather than vice versa. A container or landmark with the wrong properties in the right place is taken to be the correct container or landmark, while a container or landmark with the correct properties in the wrong place is taken to be a different container or different landmark.
From the above, it is clear that COGNITIVE MAPS are a critical component in animal navigation, just as conventional maps are in conventional navigation. A cognitive map is a representation of the layout of the environment. Its properties and the process by which it is constructed are questions of basic importance. These cognitive maps are known to be Euclidean, sense-preserving representations of the environment, that is, they encode the metric relations (distances and angles) and the sense relation (right versus left). This is most readily demonstrated by showing an animal the location of a hidden goal within a featureless rectangular room, then inertially disorienting the animal (by slow rotation in the dark) before allowing it to search for the goal. Animals of diverse species search principally at the correct location and its rotational equivalent, for example, in the correct corner and the corner diagonally opposite to it (Cheng 1986; Hermer and Spelke 1996; Margules and Gallistel 1988). They rarely look elsewhere, for example, in the other two corners. To distinguish between the diagonals of a rectangle, however, they must record both metric relations and sense relations on their map of the rectangular room. The short wall is on the left for one diagonal, regardless of which way one faces along that diagonal, but on the right for the other diagonal. If an animal could not distinguish walls based on their length (a metric relationship), or on which was to the left and which to the right (a sense relationship), then it could not distinguish between the diagonals of a rectangle.
Until recently, there were no suggestions about how animals might construct a Euclidean map of their environment. They only perceive small portions of it at any one time, so how can they perceive the relations between these portions? Recently, it has been suggested that the dead reckoning process is the key to the construction of the map (Gallistel 1990; Gallistel and Cramer 1996; McNaughton et al. 1996) because it specifies the Euclidean relationship between different points of view within a geocentric framework, a system of coordinates anchored to the earth, as opposed to an egocentric framework, a system of coordinates anchored to the animal's body (egocentric position vector in figure). Rotating the egocentric position vector by the animal's heading (its orientation in the geocentric framework) and adding the rotated vector to the geocentric position vector provided by the dead reckoning process carries the representation of the perceived portion of the environment into a common geocentric positional framework (landmark's geocentric position vector in figure). This method of map construction, in which the animal's dead reckoning automatically represents its position on its cognitive map, tells us much about how the animal perceives the shape of its environment.
Carr, H., and J. B. Watson. (1908). Orientation of the white rat. Journal of Comparative Neurology and Psychology 18:27-44.
Cheng, K. (1986). A purely geometric module in the rat's spatial representation. Cognition 23:149-178.
Collett, T. S., and A. Kelber. (1988). The retrieval of visuo-spatial memories by honeybees. Journal of Comparative Physiology, series A 163:145-150.
Gallistel, C. R. (1990). The Organization of Learning. Cambridge, MA: MIT Press.
Gallistel, C. R., and A. E. Cramer. (1996). Computations on metric maps in mammals: Getting oriented and choosing a multi-destination route. Journal of Experimental Biology 199:211-217.
Hermer, L., and E. Spelke. (1996). Modularity and development: The case of spatial reorientation. Cognition 61:195-232.
Margules, J., and C. R. Gallistel. (1988). Heading in the rat: Determination by environmental shape. Animal Learning and Behavior 16:404-410.
McNaughton, B. L., C. A. Barnes, J. L. Gerrard, K. Gothard, M. W. Jung, J. J. Knierim, H. Kudrimoti, Y. Qin, W. E. Skaggs, M. Suster, and K. L. Weaver. (1996). Deciphering the hippocampal polyglot: The hippocampus as a path integration system. Journal of Experimental Biology 199:173-185.
Neuweiler, G., and F. P. Möhres. (1967). Die Rolle des Ortgedächtnisses bei der Orientierung der Grossblatt-Fledermaus Megaderma lyra. Zeitschrift für vergleichende Physiologie 57:147-171.
Stoltz, S. P., and D. F. Lott. (1964). Establishment in rats of a persistent response producing net loss of reinforcement. Journal of Comparative and Physiological Psychology 57:147-149.
Tinkelpaugh, O. L. (1932). Multiple delayed reaction with chimpanzee and monkeys. Journal of Comparative Psychology 13:207-243.
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology