Roger Wolcott Sperry (1913-1994), who received the Nobel Prize in Physiology and Medicine in 1981, made pathfinding contributions to the sciences of brain and mind through half a century. His experiments on nerve regeneration, on cortical mechanisms of perception and learning in split-brain cats and monkeys, and on hemispheric modes of consciousness in commissurotomy patients, display outstanding creativity and skill. The intrinsic factors of nerve net patterning and in psychological action and awareness that Sperry discovered are fundamental to cognitive science.
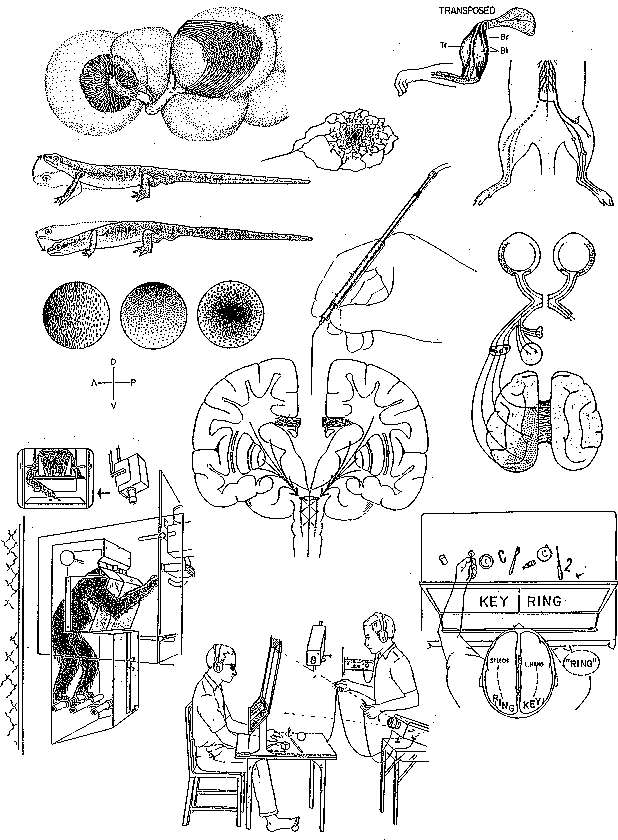
Figure 1 Selected drawings from articles by Roger Sperry, illustrating phases of his scientific career.
Mastery of his scientific vocation came quickly for Roger Sperry. While an english literature major at Oberlin, he chose a masters project with R. H. Stetson, an expert on timing in speech and music. Sperry's first paper on this myographic analysis of arm movements appeared in 1939. He proved that each time the subject repeated the same circle in space, electrical activity appeared in different muscles. The constancy of movement implied an internal image of the circular act. That year, with Paul Weiss at Chicago, Sperry published his Ph.D. abstract on the behavioral effects of surgical transposition of hindlimb muscles in rats. Within three years he had tested motor nerve regeneration in rats and monkeys, noting important species differences in motor plasticity. He also began experiments with newts that brought international renown, on growth of eye-to-brain connections after surgical rotations of the eyes (Evarts 1990). Results proved that chemical markers, previously envisaged by developmental studies of Ramon y CAJAL, Harrison, and Langley, were guiding prefunctional visual projections, even when rearranged eye-to-body relations produced totally nonfunctional reactions to stimuli (Hunt and Cowan 1990). This radically qualified prevailing theories of the construction of functional brain circuits by learning.
Experiments with optic nerve regeneration in fish after eye rotation uncovered a central patterning of movements that Sperry (1950) named "corollary discharge from efference." Independently, von Holst and Mittelstaedt in Germany had seen the same phenomenon, labeling it the "reafference principle." This proved a neural basis for the psychological fact that formulation of a motor purpose transforms processing of perceptual information. Sperry and the German scientists fully appreciated its importance in explaining the perceptual constancies, and the stability of the world seen by a moving eye. Current motor imagery theory identifies intrinsic processes defining the space-time frame for experience (Jeannerod 1994). Sperry's motor theory of perception made the same point.
In an influential paper, "Neurology and the Mind-Brain Problem," Sperry took the position that preoccupation with input to perception and cognition, disregarding constraints imposed by prewired motives or prospective MOTOR CONTROL, often leads psychology into arcane and unprofitable theorizing (Sperry 1952). His concept of biological determination of functional perceptual-motor systems met with resistance from the behaviorist psychology of his day. How he overcame this, and the strength of his evidence for chemical guidance of regenerating nerve fibers, has been reviewed recently (Evarts 1990; Levi-Montalcini 1990; Hunt and Cowan 1990). Now Sperry's principle of "chemo-affinity" as a constraining factor mapping both nerve cell interactions in the embryo and nerve tract regeneration in lower vertebrates (Sperry 1963) is beyond contention, despite powerful abstract theories developed to model order emerging in complex nonlinear dynamic systems of nerve cells, axonal and dendritic branches, and synapses, and evidence for selective retention of these elements under the validation of environmental input. Emergent order and selection create new functions, but these carry the imprint of the constraints of cell-to-cell communication set up prefunctionally in the embryo.
Surgical skills that he developed in operating on small fish enabled Sperry to attempt direct interference with corticocortical circuits in the cat by delicate subpial slicing of white matter to test various "field" theories of form perception. He proved that high-acuity vision of form must involve fiber loops leaving and returning to the cortex. Then, his interest in intrinsic whole-brain function in awareness led, by elegant logic, to division of the corpus callosum, the major axonal bridge between cortical circuits in the two cerebral hemispheres that mediate ideation and conscious guidance of purposes. Split-brain studies of cats and monkeys demonstrated that awareness and learning could be surgically bisected. They also showed that the aim of attention and voluntary responses around the subject's body, and their evolution in time, invoked a great array of structures, including undivided brain stem mechanisms (Sperry 1961).
Later, with the clinical application of commissurotomy to treat multifocal epilepsy, the split-brain approach was extended to human patients, opening the way to tests of active rational consciousnesses in left and right hemispheres, their convergent influence over acts and ideas of the whole person, and the role of language (Sperry, Gazzaniga, and Bogen 1969; Levy 1990). This research brought dramatic scientific confirmation of Sperry's theory of the causal potency of conscious ideas and beliefs. Commissurotomy research at CalTech boosted neuropsychological analysis of hemispheric differences, and, by clarifying the special modes of processing in isolated cortical territories, prompted the "cognitive revolution" of the 1960s (Sperry 1993).
In his scientific work Sperry had an idealist impulse that led him beyond searching for motive principles in awareness. In the 1960s, he formulated a philosophy of natural humanistic values, and enunciated ethical principles that recognize the innate causal power of human consciousness, for both good and ill (Sperry 1965, 1983). In the last ten years, 95 percent of his publications were on these more philosophical matters. Sperry claimed that his theory of "downward causation" or "macro-determinism" gave a new paradigm for resolving tensions between science and religion. At first, scientific colleagues thought he strayed from objectivity, but now it is clear that he foresaw the need for a larger perspective on brain activities at their highest level of organization, in human purposes, ideas, and beliefs. The foray into ethics and issues of values in policy and practice now seems prophetic. It is now far from strange to insist that a science of values is necessary.
Sperry's achievements depended on skills he showed in his earliest research -- incisive anatomical logic, imagination for the inner dynamics of psychological action, and a gift for clear writing. Sperry's psychobiology was perfectly balanced between the hard anatomical and physiological facts of the brain in communication with the freely moving body, and an imaginative, creative psychology of the mind, with all of its ethical implications. This balance he described as a "monist" position, mind and matter as inseparable parts of natural psychological processes, and he worked tirelessly to explain his view of the continuity between everyday phenomena of consciousness and the matter of the brain that scientists observe.
Evarts, E. V. (1990). Foreword: Coordination of movement as a key to higher brain function: Roger W. Sperry's contributions from 1939 to 1952. In C. Trevarthen, Ed., Brain Circuits and Functions of the Mind: Essays in Honor of Roger W. Sperry. New York: Cambridge University Press, pp. xiii - xxvi.
Hunt, R. K., and W. M. Cowan. (1990). The chemoaffinity hypothesis: An appreciation of Roger W. Sperry's contributions to developmental biology. In C. Trevarthen, Ed., Brain Circuits and Functions of the Mind: Essays in Honor of Roger W. Sperry. New York: Cambridge University Press, pp. 19-74.
Jeannerod, M. (1994). The representing brain: Neural correlates of motor intention and imagery. Behavioral and Brain Sciences 17:187-245.
Levi-Montalcini, R. (1990). Ontogenesis of neural nets: the chemoaffinity theory, 1963-1983. In C. Trevarthen, Ed., Brain Circuits and Functions of the Mind: Essays in Honor of Roger W. Sperry. New York: Cambridge University Press, pp. 3 - 18.
Levy, J. (1990). Regulation and generation of perception in the asymmetric brain. In C. Trevarthen, Ed., Brain Circuits and Functions of the Mind: Essays in Honor of Roger W. Sperry. New York: Cambridge University Press, pp. 231-248.
Sperry, R. W. (1950). Neural basis of the spontaneous optokinetic response produced by visual inversion. Journal of Comparative and Physiological Psychology 43:483-489.
Sperry, R. W. (1952). Neurology and the mind-brain problem. American Scientist 40:291-312.
Sperry, R. W. (1961). Cerebral organization and behavior. Science 133:1749-1757.
Sperry, R. W. (1963). Chemoaffinity in the orderly growth of nerve fiber patterns and connections. Proceedings of the National Academy of Sciences 50:703-710.
Sperry, R. W. (1965). Mind, brain and humanist values. In J. R. Platt, Ed., New Views on the Nature of Man. Chicago: University of Chicago Press, pp. 71-92.
Sperry, R. W. (1983). Science and Moral Priority. New York: Columbia University Press.
Sperry, R. W. (1993). The impact and promise of the cognitive revolution. American Psychologist 48(3):878-885.
Sperry, R. W., M. S. Gazzaniga, and J. E. Bogen. (1969). Interhemispheric relationships: The neocortical commissures; syndromes of hemisphere disconnection. In P. J. Vinken, and G. W. Bruyn, Eds., Handbook of Clinical Neurology, vol. 4. Amsterdam: North Holland, pp. 273-290.
Sperry, R. W. (1945). The problem of central nervous reorganization after nerve regeneration and muscle transposition. Quarterly Review of Biology 20:311-369.
Sperry, R. W. (1958). Physiological plasticity and brain circuit theory. In H. F. Harlow and C. N. Woolsey, Eds., Biological and Biochemical Bases of Behavior. Madison: University of Wisconsin Press, pp. 401-421.
Sperry, R. W. (1959). The growth of nerve circuits. Scientific American 201:68-75.
Sperry, R. W. (1964). Problems outstanding in the evolution of brain function: James Arthur lecture on the evolution of the human brain. In R. Duncan and M. Weston-Smith, Eds., American Museum of Natural History and the Encyclopedia of Ignorance. Oxford: Pergamon Press.
Sperry, R. W. (1964). The great cerebral commissure. Scientific American 210:42-52.
Sperry, R. W. (1965). Embryogenesis of behavioral nerve nets. In R. L. Dehaan and H. Ursprung, Eds., Organogenesis. New York: Holt, Rinehart and Winston, pp. 161-185.
Sperry, R. W. (1968). Mental unity following surgical disconnection of the cerebral hemispheres. The Harvey Lectures, 1966-1967. (Series 62). New York: Academic Press, pp. 293 - 323.
Sperry, R. W. (1969). A modified concept of consciousness. Psychological Reviews 76:532-536.
Sperry, R. W. (1970). Perception in absence of the neocortical commissures. Research Publications of the Association for Research in Nervous and Mental Diseases 48:123-138.
Sperry, R. W. (1974). Lateral specialization in the surgically separated hemispheres. In F. Schmitt and F. Worden, Eds., The Neurosciences. Third Study Program. Cambridge, MA: MIT Press, pp. 5-19.
Sperry, R. W. (1974). Science and the problem of values. Zygon 9:7-21.
Sperry, R. W. (1977). Forebrain commissurotomy and conscious awareness. Journal of Medical Philosophy 2(2):101-126.
Sperry, R. W. (1982). Some effects of disconnecting the cerebral hemispheres. Nobel lecture. Science 217:1223-1226.
Sperry, R. W. (1984). Consciousness, personal identity and the divided brain. Neuropsychologia 22(6):661-673.
Sperry, R. W. (1987). Consciousness and causality. In R. L. Gregory, Ed., Oxford Companion to the Mind. Oxford: Oxford University Press, pp. 164-166.
Sperry, R. W. (1988). Psychology's mentalist paradigm and the religion/science tension. American Psychologist 43(8):607-613.
Sperry, R. W. (1992). Paradigms of belief, theory and metatheory. Zygon 27:245-259 .
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology