The retina is a sheet of brain tissue lining the rear of the eye. It transduces light imaged by the lens into electrical potentials, performs substantial processing on these signals, and transmits them to the brain. This initiates visual perception.
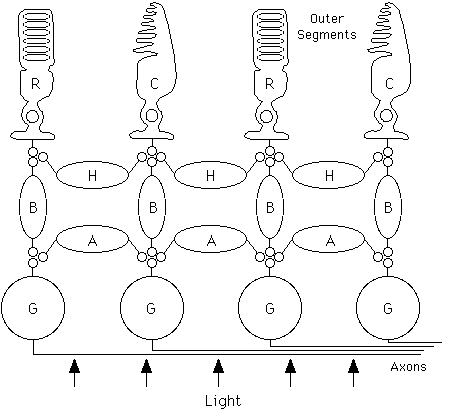
Less than half a millimeter thick, the retina consists of five major classes of neurons in three layers (see figure 1) as well as Müller glial cells. Rod and cone photoreceptor cells use an enzymatic amplification cascade to transduce light into graded electrical potentials. Rods (100 million per eye) mediate night vision and can reliably signal the absorption of an individual photon. Cones (5 million per eye) mediate daylight vision and adaptively adjust their sensitivity to register intensity over the extraordinary range of eight orders of magnitude. Rod and cone signals are transmitted to bipolar and horizontal cells which, along with amacrine cells, further process visual information with graded electrical potentials. Retinal ganglion cells integrate inputs from bipolar and amacrine cells and send the resulting visual signals as discrete action potentials (see NEURON) along 1.2 million nerve fibers to a number of targets in the brain.
Figure 1 Schematic of retinal neurons and major connections. Light traverses the mostly transparent retina before being absorbed in the outer segments of the (R)od and (C)one photoreceptors (drawn schematically) where transduction takes place. (H)orizontal, (B)ipolar and (A)macrine cells further process visual signals. (G)anglion cells integrate information from bipolar and amacrine cells and send the resulting signals to the brain via long axons.
Each of these major classes of neurons can be further divided into subclasses defined by distinctive morphology, connectivity, electrical responses to light, and biochemistry (see Rodieck 1988, 1998; Wässle and Boycott 1991). For example, there are four major subclasses of photoreceptors (rods and three types of cones) and at least nine types of ganglion cells. Though the major classes of neurons appear in all vertebrates, the characteristics of the subclasses vary considerably among species. Each known subclass is distributed across the retina, forming a mosaic that samples the visual world. Cones and many other neurons are concentrated in and near the fovea, the area of the retina corresponding to the center of gaze; here visual acuity is highest. Rods are absent in the fovea and concentrated in the peripheral retina.
Retinal ganglion cells provide visual information to the brain in signals that reflect spatial integration and parallel processing in the retina (see Rodieck 1973; Dowling 1987; Kuffler, Nichols, and Martin 1984; Davson 1972; Wandell 1995). A ganglion cell typically receives inputs over a roughly circular region of the visual field sampled by many photoreceptors. This region, the receptive field, usually consists of two concentric parts: a center region and a larger antagonistic surround region. In ON-center cells an increase in light intensity in the center increases the frequency of action potentials sent to the brain, while an increase in light intensity in the surround reduces action potential frequency. In OFF-center cells the effects of center and surround are reversed. This ON/OFF polarity is conferred by inputs from distinct types of bipolar cells which initiate the ON/OFF dichotomy (in photoreceptors the light response has a single polarity).
The ON and OFF pathways are examples of retinal subcircuits that carry different types of visual information. Likewise, rod and cone signals are processed by distinct types of bipolar and amacrine cells before being integrated in retinal ganglion cells. The subcircuits that give rise to other differences in the light responses of ganglion cells are less well understood. For example, some types of ganglion cells respond transiently at the onset or offset of a step of light while others respond for the duration of the step. Different types of ganglion cells also exhibit different spectral sensitivities determined by the relative strength of input from the three types of cones. As more subtle anatomical and physiological subdivisions of retinal neurons and their interconnections are elucidated (Rodieck 1988, 1998; Wässle and Boycott 1991), it is becoming clear that different visual messages are carried by a system of parallel retinal subcircuits. These messages are typically transmitted along different anatomical pathways to the brain where they may be locally processed in specific functional modules (see VISUAL PROCESSING STREAMS).
The operation of the retina limits visual performance. Spatial acuity is limited by the density of rods and cones and perhaps by spatial integration in the ganglion cells. Sensitivity to very dim lights is limited by the capacity of rods to faithfully signal absorption of single photons and by the capacity of subsquent circuits to transmit these signals uncorrupted by noise. Sensitivity to rapid variations in light intensity is limited by temporal integration in rods and cones (which occurs over intervals of about 300 msec and 50 msec respectively).
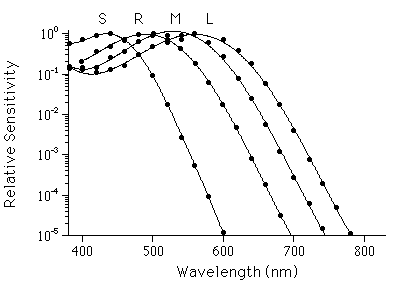
Figure 2 Photoreceptor spectral sensitivities. Points inidicate electrophysiologically measured relative spectral sensitivities of (R)ods and (L)ong, (M)iddle, and (S)hort wavelength-sensitive cones of the monkey, Macaca fascicularis, with peak sensitivities at about 500 nm, 561 nm, 531 nm, and 430 nm respectively. Human photoreceptors have very similar spectral sensitivities. Smooth curves are drawn through points to aid visualization. (Replotted from Baylor, Nunn, and Schnapf 1984, 1987.)
The operation of the rods and cones also establishes quantitative characteristics of color perception. Transduction in rods is initiated by a single visual pigment, rhodopsin. Rhodopsin has a characteristic probability of absorbing photons of different wavelength, giving the rod a characteristic spectral sensitivity (figure 2). However, once absorbed, all photons have the same effect on rhodopsin, so a rod cannot distinguish between one incident photon of 500-nm light and fifty-seven incident photons of 600-nm light. This property, called univariance, means that intensity and wavelength information are confounded in rod vision. Perceptual experience reflects this limitation: at night the world appears in shades of gray. Similarly, each of the three major cone types has a characteristic spectral sensitivity endowed by its visual pigment. Quantum absorptions in the three types of cones (figure 2) provide the visual system with just three distinct components of information about the wavelength composition of light. Hence human color perception is limited to sensations that can be duplicated by the superposition of three primary lights.
Why is wavelength information so poorly represented? The answer may lie in a compromise between spatial and spectral resolution. Each point in the visual field is sampled by only one photoreceptor. Additional types of photoreceptors might improve spectral resolution, but only at the cost of spatial resolution within each spectral band. Such trade-offs have undoubtedly had a significant impact on the evolution of retinal structure and function.
Baylor, D. A., B. J. Nunn, and J. L. Schnapf. (1984). The photo-current, noise and spectral sensitivity of rods of the monkey Macaca fascicularis.J. Physiol. (Lond.) 357:575-607.
Baylor, D. A., B. J. Nunn, and J. L. Schnapf. (1987). Spectral sensitivity of cones of the monkey Macaca fascicularis.J. Physiol. (Lond.) 390:145-160.
Davson, H. (1972). Physiology of the Eye. New York: Academic Press.
Dowling, J. E. (1987). The Retina: An Approachable Part of the Brain. Cambridge, MA: Harvard University Press.
Kuffler, S. W., J. G. Nicholls, and A. R. Martin. (1984). From Neuron to Brain. Sunderland, MA: Sinauer.
Rodieck, R. W. (1973). The Vertebrate Retina. San Francisco: W. H. Freeman.
Rodieck, R. W. (1988). The primate retina. Comparative Primate Biology 4:203-278.
Rodieck, R. W. (1998). The First Steps in Seeing. Sunderland, MA: Sinauer.
Wandell, B. A. (1995). Foundations of Vision. Sunderland, MA: Sinauer.
Wässle, H., and B. B. Boycott. (1991). Functional architecture of the mammalian retina. Physiol. Rev. 71:447-480.
Spillman, L., and J. S. Werner, Eds. (1990). Visual Perception: The Neurophysiological Foundations. San Diego: Academic Press .
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology