Columns and Modules
The CEREBRAL CORTEX sits atop the white matter of the brain, its 2
mm thickness subdivided into about six layers. Neurons with similar
interests tend to cluster. Columns are usually subdivisions at the
submillimeter scale, and modules are thought to occupy the intermediate
millimeter scale, between maps and columns. Empirically, a column
is simply a submillimeter region where many (but not all) neurons
seem to have functional properties in common. They come in two sizes,
with separate organizational principles. Minicolumns are about 23-65
µm across, and there are hundreds of them
inside any given 0.4 - 1.0 mm macrocolumn.
Each
cerebral hemisphere has about 52 "areas" distinguished
on the basis of differences between the thickness of their layers;
on average, a human cortical area is about half the size of a business
card. Though area 17 seems to be a consistent functional unit, other
areas prove to contain a half-dozen distinct physiological subdivisions
("maps") on the centimeter scale.
Both columns and modules may be regarded
as the outcomes of a self-organizing tendency during development,
patterns that emerge as surely as the hexagons of a honeycomb arise from
the pounding of so many hemispherical bee's heads on the
soft wax of tunnel walls. The hexagonal shape is an emergent property
of such competition for territory; similar competition in the cortex
may continue throughout adult life, maintaining a shifting mosaic
of cortical columns.
A column functionally ties together
all six layers. Layers II and III can usually
be lumped together, as when one talks of the superficial pyramidal
neurons. But layer IV has had to be repeatedly subdivided
in the visual cortex (IVa, IVb, IVc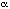 , IVc
, IVc
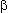 ). Layer IV neurons send most of their
outputs up to II and III. Some superficial neurons
send messages down to V and VI, though their most prominent
connections (either laterally in the layers or via U-fibers in white
matter) are within their own layers. Layer VI sends messages
back down to the THALAMUS via the white matter, while V sends
signals to other deep and distant neural structures, sometimes even
the spinal cord.
). Layer IV neurons send most of their
outputs up to II and III. Some superficial neurons
send messages down to V and VI, though their most prominent
connections (either laterally in the layers or via U-fibers in white
matter) are within their own layers. Layer VI sends messages
back down to the THALAMUS via the white matter, while V sends
signals to other deep and distant neural structures, sometimes even
the spinal cord.
For any column of cortex, the bottom
layers are like a subcortical outgoing mailbox, the middle layer
like an inbox, and the superficial layers somewhat like an interoffice
mailbox spanning the columns and reaching out to other cortical areas
(Calvin and Ojemann 1994). Indeed, Diamond (1979) argues that the "motor
cortex" isn"t restricted to the motor strip but
is the fifth layer of the entire cerebral cortex. That"s because V,
whatever the area, contains neurons that at some stage of development
send their outputs down to the spinal cord, with copies to the brain
stem, BASAL GANGLIA, and hypothalamus. Likewise
the fourth layer everywhere is the "sensory cortex," and
the second and third layers everywhere are the true "association
cortex" in this view.
Minicolumns appear to be formed from
dendritic bundles. Ramón y CAJAL saw
connect-the-dots clusters of cell bodies, running from white matter
to the cortical surface; these hair-thin columns are about 30 m
apart in human cortex. It now appears that a column is like a stalk
of celery, a vertical bundle containing axons and apical dendrites
from about 100 neurons (Peters and Yilmaz 1993) and their internal
microcircuitry.
Macrocolumns may, in contrast, reflect
an organization of the input wiring, for example, corticocortical
terminations from different areas often terminate in interdigitating
zones about the width of a thin pencil lead. In 1957, Mountcastle
and his coworkers discovered a tendency for somatosensory strip neurons
responsive to skin stimulation (hair, light touch) to alternate
with those specializing in joint and muscle receptors about every
0.5 mm. It now appears that there is a mosaic organization of similar
dimensions, the neurons within each macrocolumn (or "segregate")
having a receptive field optimized for the same patch of body surface (Favorov
and Kelly 1994).
Hubel and Wiesel, recording in monkey
visual cortex, saw curtainlike clusters ("ocular dominance
columns") that specialized in the left eye, with an adjacent
cluster about 0.4 mm away specializing in the right eye. As seems
appropriate for an outcome of self-organization, average size varies
among individuals over 0.4-0.7 mm; those with smaller ocular dominance
columns have more of them (Horton and Hocking 1996).
Orientation columns are
of minicolumn dimensions, within which the neurons prefer lines
and edges that are tilted about the same angle from the vertical;
there are many such minicolumns specializing in various angles within
an ocular dominance macrocolumn (Hubel and Wiesel 1977). The relationships
between minicolumns and macrocolumns are best seen in VISUAL CORTEX, though it may be hazardous to generalize from
this because ocular dominance columns themselves are less than universal;
for instance, they are not a typical feature of New World monkeys.
Color blobs are clusters
of COLOR-sensitive neurons in the cortex at
macrocolumnar spacing but involving only neurons of the superficial
layers and not extending throughout all cortical layers, as in a
proper column. Recently, recurrent excitation in the superficial
layers has been identified as a coordinating (and perhaps self-organizing)
principle among distant minicolumns (Calvin 1995). The superficial
pyramids send myelinated axons out of the cortical layers into the
white matter; their eventual targets are typically the superficial
layers of other cortical areas when of the "feedback" type;
when "feedforward" they terminate in IV and
deep III.
But superficial pyramidal neurons
also send unmyelinated collaterals sideways, with an unusual patterning
that suggests a columnar organizing principle. Like an express train
that skips intermediate stops, the collateral axon travels a characteristic
lateral distance without giving off any terminal branches; then
it produces a tight terminal cluster (see fig. 5 in Gilbert 1993).
The axon may continue for many millimeters, repeating such clusters
about every 0.43 mm in primary visual cortex, 0.65 mm in the secondary
visual areas, 0.73 mm in sensory strip, and 0.85 mm in motor cortex
of monkeys (Lund, Yoshioka, and Levitt 1993). Because of this local
standard for axon length, mutual re-excitation becomes probable
among some cell pairs. Macrocolumns of similar emphasis are seen
to be connected by such synchronizing excitation. Calvin (1996)
argues that these express connections could implement a Darwinian
copying competition among Hebbian cell-assemblies on the time scale
of thought and action, providing one aspect of CONSCIOUSNESS.
Though COMPUTATIONAL NEUROANATOMY has proved
more complex, it has been widely expected that cerebral cortex will
turn out to have circuits which, in different cortical patches,
are merely repeats of a standard "modular" pattern,
something like modular kitchen cabinets. Columns, barrels, blobs,
and stripes have all been called modules, and the term is loosely
applied to any segmentation or repeated patchiness (Purves, Riddle,
and LaMantia 1992) and to a wide range of functional or anatomical
collectives. The best candidate for a true module was the "hypercolumn" (Hubel
and Wiesel 1977): two adjacent ocular dominance columns, each containing
a full set of orientation columns, suggested similar internal wiring,
whatever the patch of visual field being represented. However, newer mapping
techniques have shown that ocular dominance repeats are somewhat
independent of orientation column repeats (Blasdel 1992), making
adjacent hypercolumns internally nonidentical, that is, not iterated
circuitry. Module remains a fuzzy term for anything larger than
a macrocolumn but smaller than a map -- though one increasingly
sees it used as a trendy word denoting any cortical specialization, for
example, modules as the foundation for "multiple intelligences."
See also
Additional links
-- William H. Calvin
References
Bartfeld, E., and A. Grinvald. (1992). Relationships
between orientation -preference pinwheels, cytochrome oxidase
blobs, and ocular-dominance columns in primate striate cortex. Proc.
Natl. Acad. Sci. USA 89:11905-11909.
Blasdel, G. G. (1992). Orientation selectivity,
preference, and continuity in monkey striate cortex. J. Neurosci. 12:3139-3161.
Bullock, T. H. (1980). Reassessment of neural
connectivity and its specification. In H. M. Pinsker and W. D. Willis,
Jr., Eds., Information Processing in the Nervous System. New
York: Raven Press, pp. 199-220.
Calvin, W. H. (1995). Cortical columns, modules,
and Hebbian cell assemblies. In Michael A. Arbib, Ed., The
Handbook of Brain Theory and Neural Networks. Cambridge,
MA: Bradford Books/MIT Press, pp. 269-272.
Calvin, W. H. (1996). The Cerebral Code:
Thinking a Thought in the Mosaics of the Mind. Cambridge,
MA: MIT Press.
Calvin, W. H., and G. A. Ojemann. (1994). Conversations
with Neil"s Brain: The Neural Nature of Thought and Language. Reading,
MA: Addison-Wesley.
Diamond, I. (1979). The subdivisions of neocortex:
A proposal to revise the traditional view of sensory, motor, and
association areas. In J. M. Sprague and A. N. Epstein, Eds., Progress
in Psychobiology and Physiological Psychology 8. New York:
Academic Press, pp. 1-43.
Favorov, O. V., and D. G. Kelly. (1994). Minicolumnar
organization within somatosensory cortical segregates: I. Development
of afferent connections. Cerebral Cortex 4:408-427.
Gilbert, C. D. (1993). Circuitry, architecture,
and functional dynamics of visual cortex. Cerebral Cortex 3:373-386.
Goldman-Rakic, P. (1990). Parallel systems in
the cerebral cortex: The topography of cognition. In M. A. Arbib
and J. A. Robinson, Eds., Natural and Artificial Parallel
Computation. Cambridge, MA: MIT Press, pp.155-176.
Horton, J. C., and D. R. Hocking. (1996). Intrinsic
variability of ocular dominance column periodicity in normal macaque
monkeys. J. Neurosci. 16(22):7228-7239.
Hubel, D. H., and T. N. Wiesel. (1977). Functional
architecture of macaque visual cortex. Proc. Roy. Soc. (London) 198B:
1-59.
Katz, L. C., and E. M. Callaway. (1992). Development
of local circuits in mammalian visual cortex. Ann. Rev. Neurosci. 15:31-56.
Livingstone, M. S. (1996). Oscillatory firing
and interneuronal correlations in squirrel monkey striate cortex. J.
Neurophysiol. 75:2467-2485.
Livingstone, M. S., and D. H. Hubel. (1988).
Segregation of form, color, movement, and depth: Anatomy, physiology,
and perception. Science 240:740-749.
Lund, J. S., T. Yoshioka, and J. B. Levitt.
(1993). Comparison of intrinsic connectivity in different areas
of macaque monkey cerebral cortex. Cerebral Cortex 3:148-162.
Mountcastle, V. B. (1979). An organizing principle
for cerebral function: The unit module and the distributed system.
In F. O. Schmitt and F. G. Worden, Eds., The Neurosciences
Fourth Study Program. Cambridge, MA: MIT Press, pp. 21-42.
Peters, A., and E. Yilmaz. (1993). Neuronal
organization in area 17 of cat visual cortex. Cerebral Cortex 3:49-68.
Purves, D., D. R. Riddle, and A-S. LaMantia.
(1992). Iterated patterns of brain circuitry (or how the cortex
gets its spots). Trends in the Neurosciences 15:362-368
(see letters in 16: 178 - 181).
Shaw, G. L., E. Harth, and A. B. Scheibel. (1982).
Cooperativity in brain function: Assemblies of approximately 30
neurons. Exp. Neurol. 77:324-358.
White, E. L. (1989). Cortical Circuits. Boston: Birkhauser.
Yuste, R., and D. Simons. (1996). Barrels in
the desert: the Sde Boker workshop on neocortical circuits. Neuron 19:231-237 .
 Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology