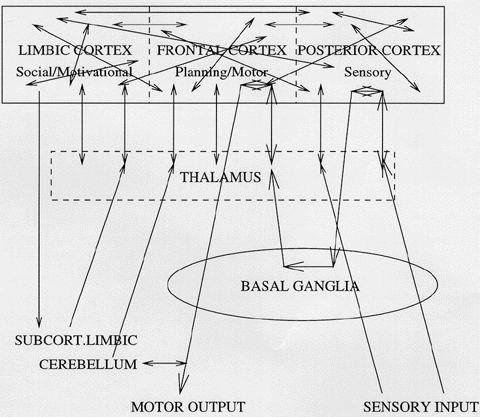
Figure 1 A simplified schematic of the architecture of mammalian brains, indicating the main connections of the thalamus.
The thalamus is a component of the brain of every mammal: it looks like a pair of small eggs, one underneath each of the cerebral hemispheres. Its volume is 2 percent and 4 percent of the volume of the cortex. It has a simple position in the overall architecture: virtually all information arriving at the CEREBRAL CORTEX comes from the thalamus, which receives it from subcortical structures. This architecture is shown in figure 1. In particular, all visual, auditory, tactile, and proprioceptive information passes through the thalamus on its way to cortex; all planning and motor information generated by the CEREBELLUM or by the BASAL GANGLIA is passed through the thalamus to cortex; and emotional or motivational data from the AMYGDALA and the mammilary body passes through the thalamus also. The main exceptions to this are (1) the direct pathway from the olfactory bulb to olfactory cortex; (2) a supplementary direct pathway from the amygdala to prefrontal cortex; and (3) diffuse neuromodulatory projections from various brain stem nuclei to cortex. These facts give rise to the classic view that the thalamus is a passive relay station which generates virtually all the information bearing input to the cortex. This view is strengthened by the simplicity of the internal connections in the thalamus: the pathways above are built from a homogeneous population of excitatory "relay" cells which give off almost no local collaterals in the thalamus, but synapse strongly on layer IV (and deep layer III) cortical neurons. Finally, this "feedforward" pathway is topographically organized, with various nuclei of the thalamus projecting to various areas of cortex and a "rod-to-column" pattern within each nucleus (Jones 1985: fig 3.20, 3.22, and p. 811).
Figure 1 A simplified schematic of the architecture of mammalian brains, indicating the main connections of the thalamus.
BUT the above picture has omitted one fundamental fact: all projections from thalamus to cortex are reciprocated by feedback projections from cortex to thalamus of the same or even larger size. For instance, Sherman and Koch (1986) estimate that in cat there are roughly 106 fibers from the lateral geniculate nucleus in the thalamus to the VISUAL CORTEX, but 107 fibers in the reverse direction! As Jones says, "Can it be that such a highly organized and numerically dense projection has virtually no functional significance? One doubts it. The very anatomical precision speaks against it. Every dorsal thalamic nucleus receives fibers back from all cortical areas to which it projects; . . ." (Jones 1985: 819).
Feedback pathways are the hallmark of mammalian brains. Lower animals have brains which are, by and large, constructed on a feedforward, modular architecture. In the total reorganization that formed the mammalian telencephalon, two vast families of feedback pathways appear: the corticothalamic pathways just described and the corticocortical feedback pathways, by which virtually all nonlocal connections within the cortex (the myelinated tracts in the white matter connecting distinct cortical areas) are reciprocated. This architecture is also indicated schematically in figure 1. From an information-processing point of view, modeling the role of these feedback pathways seems to be one of the first steps in making sense of cortical anatomy. And since the corticothalamic feedback is relatively simple, this would seem to be a prime target for models.
Perhaps the most accepted model for this feedback is that it is used to gate the information which reaches cortex, allowing the cortex to selectively attend to part of the data at a given time. Rather striking evidence for this was discovered by Singer and coworkers (Singer and Schmielau 1976; Singer 1977; Varela and Singer 1987): they found that cortical feedback was used to selectively enhance visual input from the two eyes at points where they were viewing a surface in the fixation plane. This highlights objects on the plane of fixation and suppresses objects which are nearer or farther. Subsequently Crick (1984) developed a specific mechanism for such gating, based on a small group of thalamic cells called the reticular nucleus. In his view, such thalamic gating might underlie the psychophysical results of Treisman (1980) implicating serial search in some visual tasks. The attention hypothesis has been used to explain effects in humans after thalamic strokes (Rafal and Posner 1987) and after direct electrode stimulation in the course of neurosurgery (Ojemann 1983). Very suggestive results on the enhancement and inhibition caused by coricothalamic feedback from S1 to VPM in rat have been discovered by Ghazanfar and Nicolelis (1997) .
Does mere gating require such a massive backprojection? Beautiful evidence for a subtler role of corticothalamic feedback was discovered recently by Sillito and co-workers (1994). They found that cortical feedback caused thalamic relay cells responding to different parts of a coherent visual stimulus to synchronize. More precisely, an extended moving bar excites multiple thalamic relay cells whose receptive fields lie at different positions along the bar. When cortical feedback was intact, the spike trains of these cells showed strong correlations which were absent if either the feedback was interrupted or if the stimulus was not coherent (e.g., being made up of two bars). This suggests that information can pass back and forth between thalamus and cortex and that important nonlocal patterns are recognized during this iterative operation. Further evidence is found in Nicolelis et al. (1995).
Several models for such iterative algorithms have been proposed. The earliest, to my knowledge, was the ALOPEX theory of Harth et al. (1987), described as follows: "A model is proposed in which the feedback pathways serve to modify afferent sensory stimuli in ways that enhance and complete sensory input patterns, suppress irrelevant features, and generate quasi-sensory patterns when afferent stimulation is weak or absent." I have elaborated this theory (see Mumford 1991-2, 1993), describing the role it imputes to the thalamus as that of an "active blackboard." The basic idea is that there are multiple cortical assemblies which respond to the presence of distinct nonlocal visual patterns; in a noisy ambiguous real-world stimulus, many of these will respond to varying degrees. These assemblies then compete to enhance the features they depend on while inhibiting their rival's features via their feedback on both the thalamic relay cells and the thalamic inhibitory interneurons. A related idea has been implemented in tracking algorithms in Isard and Blake (1996): their ALGORITHM infers multiple competing hypotheses which kill each other off as evidence accrues. One hopes that advances in recording techniques and the use of more realistic stimuli will enable tests of these hypotheses to be made in the coming decade.
Crick, F. (1984). Function of the thalamic reticular complex: The searchlight hypothesis. Proc. Natl. Acad. Sci., USA 81:4586-4590.
Ghazanfar, A., and M. Nicolelis. (1997). Non-linear spatiotemporal computation of tactile information by thalamocortical ensembles. J. Neurophys.78:506-510.
Harth, E., K. P. Unnikrishnan, and A. S. Pandya. (1987). The inversion of sensory processing by feedback pathways: A model of visual cognitive functions. Science 1987:184-187.
Isard, M., and A. Blake. (1996). Contour tracking by stochastic propagation of conditional density. Proc Eur. Conf. Comp. Vision 1:343-356.
Jones, E. G. (1985). The Thalamus. New York: Plenum Press.
Mumford, D. (1991-2). On the computational architecture of the neocortex: 1. The role of the thalamo-cortical loop, 2. The role of the cortico-cortical loops. Biol. Cybern. 65: 135 - 145; 66: 241 - 251.
Mumford, D. (1993). Neuronal architectures for pattern-theoretic problems. In C. Koch, Ed., Large Scale Neuronal Models of the Brain. Cambridge, MA: MIT Press.
Nicolelis, M., L. Baccala, R. Lin, and J. Chapin. (1995). Sensori-motor encoding by synchronous neural ensemble activity at multiple levels of the somoatosensory system. Science 268:1353-1358.
Ojemann, G. (1983). Brain organization for language from the perspective of electrical stimulation mapping. Behav. Brain Sci. 2:189-206.
Rafal, R., and M. Posner. (1987). Deficits in human visual attention following thalamic lesions. Proc. Natl. Acad. Sci. USA 84:7349-7353.
Sherman, M., and C. Koch. (1986). The control of retinogeniculate transmission in the mammalian LGN. Exp. Brain Res. 63:1-20.
Sillito, A., H. Jones, G. Gerstein, and D. West. (1994). Feature-linked synchronization of thalamic relay cell firing induced by feedback from the visual cortex. Nature 369:479-482.
Singer, W. (1977). Control of thalamic transmission by corticofugal and ascending reticular pathways in the visual system. Physiol. Rev. 57:386-419.
Singer, W., and F. Schmielau. (1976). The effect of reticular stimulation on binocular inhibition in the cat LGN. Exp Brain Res. 14:210-226.
Treisman, A., and G. Gelade. (1980). A feature integration theory of attention. Cognitive Psychology 12:99-136.
Varela, F., and W. Singer. (1987). Neuronal dynamics in the visual corticothalamic pathway revealed through binocular rivalry. Exp Brain Res. 66:10-20 .
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology