Electric and magnetic evoked fields are generated in the brain as a consequence of the synchronized activation of neuronal networks by external stimuli. These evoked fields may be associated with sensory, motor, or cognitive events, and hence are more generally termed event-related potentials ERPs) and event-related magnetic fields (ERFs), respectively. Both ERPs and ERFs consist of precisely timed sequences of waves or components that may be recorded noninvasively from the surface of the head to provide information about spatio-temporal patterns of brain activity associated with a wide variety of cognitive processes (Heinze, Münte, and Mangun 1994; Rugg and Coles 1995).
Electric and magnetic field recordings provide complementary information about brain function with respect to other neuroimaging methods that register changes in regional brain metabolism or blood flow, such as POSITRON EMISSION TOMOGRAPHY (PET) and functional MAGNETIC RESONANCE IMAGING (fMRI). Although PET and fMRI provide a detailed anatomical mapping of active brain regions during cognitive performance, these methods cannot track the time course of neural events with the high precision of ERP and ERF recordings. Studies that combine ERP/ERF and PET/fMRI methodologies are needed to resolve both the spatial and temporal aspects of brain activity patterns that underlie cognition.
At the level of SINGLE-NEURON RECORDING, both ERPs and ERFs are generated primarily by the flow of ionic currents across nerve cell membranes during synaptic activity. ERPs arise from summed field potentials produced by synaptic currents passing into the extracellular fluids surrounding active neurons. In contrast, ERFs are produced by the concentrated intracellular flow of synaptic currents through elongated neuronal processes such as dendrites, which gives rise to concentric magnetic fields surrounding the cells. When a sufficiently large number of neurons having a similar anatomical position and orientation are synchronously activated, their summed fields may be strong enough to be detectable as ERPs or ERFs at the surface of the head. The detailed study of scalp-recorded ERPs became possible in the 1960s following the advent of digital signal-averaging computers, whereas analysis of ERFs required the further development in the 1980s of highly sensitive, multichannel magnetic field sensors (Regan 1989).
The anatomical locations of the neuronal populations that generate ERPs and ERFs may be estimated on the basis of their surface field configurations. This requires application of algorithms and models that take into account the geometry of the generator neurons and the physical properties of the biological tissues. Active neural networks may be localized in the brain more readily by means of ERF than by surface ERP recordings, because magnetic fields pass through the brain, skull, and scalp without distortion, whereas ERPs are attenuated by the resistivity of intervening tissues. Both ERP and ERF data have been used successfully to reveal the timing of mental operations with a high degree of precision (of the order of milliseconds) and to localize brain regions that are active during sensory and perceptual processing, selective attention and discrimination, memory storage and retrieval, and language comprehension (Hillyard 1993).
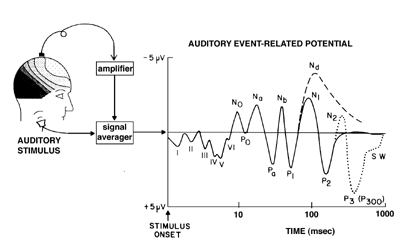
Figure 1 The characteristic time-voltage waveform of the auditory ERP in response to a brief stimulus such as a click or a tone. To extract the ERP from the ongoing noise of the electroencephalogram, it is necessary to signal average the time-locked waves over many stimulus presentations. The individual waves or components of the ERP are triggered at specific time delays or latencies after the stimulus (note logarithmic time scale). The earliest waves (I - VI) are generated in the auditory brainstem pathways, while subsequent negative (N) and positive (P) waves are generated in different subregions of primary and secondary auditory cortex. (From Hillyard, S.A. (1993). Electrical and magnetic brain recordings: contributions to cognitive neuroscience. Current Opinion in Neurobiology 3: 217-224.)
The processing of sensory information in different modalities is associated with characteristic sequences of surface-recorded ERP/ERF components (figure 1). In each modality, components at specific latencies represent evoked activity in subcortical sensory pathways and in primary and secondary receiving areas of the CEREBRAL CORTEX. Cortical components with latencies of 50-250 msec have been associated with perception of specific classes of stimuli (Allison et al. 1994) and with short-term sensory memory processes. Altered sensory experience (e.g., congenital deafness, blindness, or limb amputation) produces marked changes in ERP/ERF configurations that reflect the NEURAL PLASTICITY and functional reorganization of cortical sensory systems (Neville 1995).
Recordings of ERPs and ERFs have revealed both the timing and anatomical substrates of selective ATTENTION operations in the human brain (Näätänen 1992; Hillyard et al. 1995). In dichotic listening tasks, paying attention to sounds in one ear while ignoring sounds in the opposite ear produces a marked enhancement of short-latency ERP/ERF components to attended-ear sounds in auditory cortex. This selective modulation of attended versus unattended inputs during AUDITORY ATTENTION begins as early as 20-50 msec poststimulus, which provides strong support for theories of attention that postulate an "early selection" of stimuli prior to full perceptual analysis. In visual attention tasks, stimuli presented at attended locations in the visual field elicit enlarged ERP/ERF components in secondary (extrastriate) cortical areas as early as 80 - 100 msec poststimulus. This suggests that visual attention involves a sensory gain control or amplification mechanism that selectively modulates the flow of information through extrastriate cortex. Paying attention to nonspatial features such as color or shape is manifested by longer latency components that index the time course of feature analyses in different visual-cortical areas.
ERPs and ERFs provide a converging source of data about the timing and organization of information processing stages that intervene between a stimulus and a discriminative response. Whereas short-latency components demarcate the timing of early sensory feature analyses, longer latency components ("N200" and "P300" waves) are closely coupled with processes of perceptual discrimination, OBJECT RECOGNITION, and classification. ERP components generated in motor cortex index the timing of response selection and MOTOR CONTROL processes. Studies using these ERP measures have provided strong support for "cascade" theories that posit a continuous flow of partially analyzed information between successive processing stages during sensory-motor tasks (Coles et al. 1995).
Long-latency ERPs have been linked with MEMORY encoding, updating, and retrieval processes (Rugg 1995). ERPs elicited during LEARNING can reliably predict accuracy of recall or recognition on subsequent testing. Some components appear to index conscious recognition of previously learned items, whereas others are sensitive to contextual priming effects. These memory-related components have been recorded both from the scalp surface and from implanted electrodes in hippocampus and adjacent temporal lobe structures in neurosurgical patients.
ERP and ERF recordings are also being used effectively to investigate the NEURAL BASIS OF LANGUAGE, including phonetic, lexical, syntactic, and semantic levels of processing (Kutas and Van Petten 1994). Alterations in specific ERP/ERF components have been linked to syndromes of LANGUAGE IMPAIRMENT. A late negative ERP ("N400") provides a graded, on-line measure of word expectancy and semantic priming during sentence comprehension. Studies of N400 have contributed to understanding the organization of semantic networks in the brain (McCarthy et al. 1995).
Allison, T., G. McCarthy, A. C. Nobre, A. Puce, and A. Belger. (1994). Human extrastriate visual cortex and the perception of faces, words, numbers, and colors. Cerebral Cortex 5:544-554.
Coles, M. G. H., G. O. Henderikus, M. Smid, M. K. Scheffers, and L. J. Otten. (1995). Mental chronometry and the study of human information processing. In M. D. Rugg and M. G. H. Coles, Eds., Electrophysiology of Mind: Event-Related Brain Potentials and Cognition. Oxford: Oxford University Press, pp. 86-131.
Heinze, H. J., T. F. Münte, and G. R. Mangun, Eds. (1994). Cognitive Electrophysiology. Boston: Birkhauser.
Hillyard, S. A. (1993). Electrical and magnetic brain recordings: Contributions to cognitive neuroscience. Current Opinion in Neurobiology 3:217-224.
Hillyard, S. A., G. R. Mangun, M. G. Woldorff, and S. J. Luck. (1995). Neural systems mediating selective attention. In M. S. Gazzaniga, Ed., The Cognitive Neurosciences. Cambridge, MA: MIT Press, pp. 665-681.
Kutas, M., and C. K. Van Petten. (1994). Psycholinguistics electrified: Event-related brain potential investigations. In M. Gernsbacher, Ed., Handbook of Psycholinguistics. New York: Academic Press, pp. 83-143.
McCarthy, G., A. C. Nobre, S. Bentin, and D. D. Spence. (1995). Language-related field potentials in the anterior-medial temporal lobe: 1. Intracranial distribution and neural generators. Journal of Neuroscience 15:1080-1089.
Näätänen, R. (1992). Attention and Brain Function. Hillsdale, NJ: Erlbaum.
Neville, H. (1995). Developmental specificity in neurocognitive development in humans. In M. S. Gazzaniga, Ed., The Cognitive Neurosciences. Cambridge, MA: MIT Press, pp. 219-234.
Regan, D. (1989). Human Brain Electrophysiology. New York: Elsevier.
Rugg, M. D. (1995). ERP studies of memory. In M. D. Rugg and M. G. H. Coles, Eds., Electrophysiology of Mind: Event-Related Brain Potentials and Cognition. Oxford: Oxford University Press, pp. 132-170.
Rugg, M. D., and M. G. H. Coles, Eds. (1995). Electrophysiology of Mind: Event-Related Brain Potentials and Cognition. Oxford: Oxford University Press.
Coles, M. G. H., E. Donchin, and S. W. Porges, Eds. (1986). Psychophysiology: Systems, Processes, and Applications. Vol. 1, Systems. New York: Guilford Press.
Donchin, E., Ed. (1984). Cognitive Psychophysiology. Hillsdale, NJ: Erlbaum.
Gaillard, A. W. K., and W. Ritter, Eds. (1983). Tutorials in ERP Research: Endogenous Components. Amsterdam: North-Holland.
Hämäläinen, M., R. Hari, R. J. Ilmoniemi, J. Knuutila, and O. V. Lounasmaa. (1993). Magnetoencephalography: Theory, instrumentation, and applications to noninvasive studies of the working human brain. Reviews of Modern Physics 65:413-497.
Hillyard, S. A., L. Anllo-Vento, V. P. Clark, H. J. Heinze, S. J. Luck, and G. R. Mangun. (1996). Neuroimaging approaches to the study of visual attention: A tutorial. In M. Coles, A. Kramer, and G. Logan, Eds., Converging Operations in the Study of Visual Selective Attention. Washington, DC: American Psychological Association, pp. 107-138.
Hillyard, S., and T. W. Picton. (1987). Electrophysiology of cognition. In F. Plum, Ed., Handbook of Physiology Section 1: The Nervous System. Vol. 5, Higher Functions of the Brain. Bethesda: American Physiological Society, pp. 519-584.
John, E. R., T. Harmony, L. Prichep, M. Valdés, and P. Valdés, Eds. (1990). Machinery of the Mind. Boston: Birkhausen.
Näätänen, R. (1992). Attention and Brain Function. Hillsdale, NJ: Erlbaum.
Näätänen, R. (1995). The mismatch negativity: A powerful tool for cognitive neuroscience. Ear and Hearing 16:6-18.
Nunez, P. L., Ed. (1981). Electric Fields of the Brain. New York: Oxford University Press.
Picton, T. W., O. G. Lins, and M. Scherg. (1994). The recording and analysis of event-related potentials. In F. Boller and J. Grafman, Eds., Handbook of Neuropsychology. Vol. 9, Event-Related Potentials. Amsterdam: Elsevier, pp.429-499.
Scherg, M. (1990). Fundamentals of dipole source potential analysis. In F. Grandori, M. Hoke, and G. L. Romans, Eds., Auditory Evoked Magnetic Fields and Electric Potentials, Advances in Audiology. Basel: Karger, pp. 40-69 .
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology