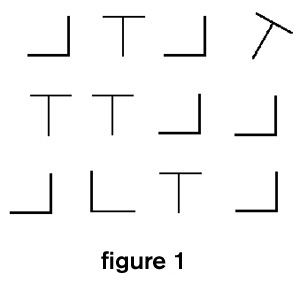
To illustrate what is meant by attention, consider the display in figure 1. Your ATTENTION may be drawn to the tilted T because it differs in such a striking way from the background. When one figure differs from the background by a single feature, it pops out and your attention is drawn to it. This is an example of attention driven by input. However, if you know that the target is an L you can guide your search among the stimuli with horizontal and vertical strokes. This is an example of the form of higher level voluntary control that is the subject of this section. Voluntary control is accompanied by the subjective feeling of selection between potential actions and is one of the most distinctive features of human experience. The interaction of top-down voluntary actions with bottom-up automatic processes, which is illustrated by figure 1, has interested researchers since the beginnings of psychology (James 1890; see WILLIAM JAMES).
One approach to the question of voluntary control is to argue that it is an illusion that arises out of the competitive activation of a large number of brain systems. What appears to be volition is the result of a complex network relaxing to a particular state. Although without denying the top-down component, this view stresses the bottom-up processes. A different view elaborated in this section is that there is a high-level executive attention network with its own anatomy that works to resolve competition and in the process gives rise to subjective feelings of cognitive control (Norman and Shallice 1986). This view emphasizes the top-down control.
The executive system participates in tasks that involve conflict between systems. This is a property one expects to find in a system that has as a major function inhibition of reflexive or bottom-up responses to external stimuli in order to allow autonomous action. A classical paradigm to study the inhibition of habitual responses is the Stroop task (Stroop 1935). In this task, subjects name the color of the ink of a word. Sometimes, the word is a color name (e.g., red) in a different ink color (e.g., blue). In those incongruent trials subjects automatically read "red" and have to inhibit this answer to respond "blue." Inhibition produces interference revealed by slow reaction times in the incongruent condition.
Is it possible to uncover the neural substrates of cognitive control? Imaging techniques developed in the last several years have yielded promising results (Toga and Mazziota 1996). An area in the medial surface of the frontal lobe, named the anterior cingulate gyrus, appears to be important for the inhibition of automatic response that is central to voluntary action. Five studies involving measurement of blood flow by positron emission tomography (PET) in a Stroop task have shown activation of the anterior cingulate in the incongruent condition when compared with the congruent condition (e.g., the noun blue displayed in blue color) or neutral (noncolor word; see Posner and DiGirolamo 1996 for a review).
Other tasks requiring inhibition of habitual responses also activate the anterior cingulate. For example, responding to a noun by generating an associated use produces more activation of the anterior cingulate than simply repeating the noun (Petersen et al. 1989). In the generate condition, the most familiar response (i.e., repeating the noun) needs to be repressed, to allow the expression of the verb. Classifying a noun into a category also produces cingulate activation related to the number of targets. This finding suggests that the anterior cingulate activation is due to special processing of the target rather than being necessary to make the classification, a result consistent with the idea of cognitive control. The cingulate has close connection to underlying subcortical areas in the BASAL GANGLIA (Houk 1995). These areas have also shown activity in some of the same tasks described above and play a role in the inhibition of reflexive motor responses. It seems likely they form part of the network subserving this form of voluntary control.
Goldberg and Bloom (1990) proposed a "dual premotor system hypothesis" of volitional movement. This theory, which attributes an executive function to the anterior cingulate and the supplementary motor area, was developed to explain the alien hand sign. The alien hand sign is the performance of apparently purposive movements that the patient fails to recognize as self-generated. The theory posits a lateral premotor system (LPS; Area 6), that organizes motor behavior in reaction to external stimulus, and a medial premotor system (MPS; anterior cingulate, supplementary motor area, and basal ganglia loops), which underlies intentional behavior. MPS underlies volitional movement by inhibiting the LPS. If a lesion occurs in MPS, LPS is released and obligatory dependence on external information emerges. The patient develops compulsive automatisms, which are not perceived as self-generated. The inhibitory effect of MPS over LPS during volitional movement resembles the inhibitory effect of MPS (i.e., anterior cingulate) over semantic networks during the Stroop task. The idea of alien rather than self control is also found in some forms of schizophrenia, a disorder that has also been shown to involve abnormalities in the anterior cingulate and basal ganglia (Benes 1993; Early 1994).
Cognitive studies have shown several forms of short term or WORKING MEMORY and considerable independence between them (Baddeley 1986). Recent imaging data show that verbal, spatial, and object memories involve separate anatomical areas (Smith and Jonides 1995). There is evidence that all forms of memory are interfaced to a common executive system that involves the same midline frontal anatomy described previously (Baddeley 1986; Posner and Raichle 1994).
PET studies have also shown that executive attention plays an important role in high level skills (Kosslyn 1994; Posner and Raichle 1994). Studies involving recording from scalp electrodes have provided some information on the time course of the activations found in PET studies during reading. Skills such as READING have a very strong dependence on rapid processing. A skilled reader fixates on a given word for only about 275 msec (Rayner and Sereno 1994). In generating the use of visual words, activation of the cingulate begins as early as 150 msec after input when blocks of trials in which subjects derive a word meaning alternate with blocks in which they read the word aloud (Snyder et al. 1995). The cingulate activation occurs whenever higher level supervisory control is needed to organize the mental response to the input. In the case of generating the use of a word, attention leads and probably is required for the activation of a network of areas that lead eventually to articulation of novel ideas associated with the input string. We see an early semantic analysis of the input word after 200 msec and development of associations to the input in frontal and parietal sites over the next second. Although it is possible to lay out a sequence of processing steps, they can be misleading. Because attention may occur rather early it is possible for subjects to reprogram the organization of these steps and thus to carry out a number of different instructions with the same brain network. Studies of the role of attention suggest that reorganization involves amplification of the operations that are attended in comparison to unattended operations. Increases in overall neuronal activity appear to produce faster speed and higher priority for the attended computations. As attention is released from high order activity during practice in the skill it becomes possible to improve the speed of performance by amplification of early processing steps.
Studies of mental arithmetic, visual IMAGERY, and other forms of skilled performance using neuroimaging methods seem to support many of the same principles that have been outlined above for word reading.
Baddeley, A. (1986). Working Memory. Oxford: Oxford University Press.
Benes, F. M. (1993). Relationship of cingulate cortex to schizophrenia and other psychiatric disorders. In B. A. Vogt and M. Gabriel, Eds., Neurobiology of Cingulate Cortex and Limbic Thalamus. Boston: Birkhauser.
Early, T. S. (1994). Left globus pallidus hyperactivity and right-sided neglect in schizophrenia. In R. L. Cromwell and C. R. Snyder, Eds., Schizophrenia: Origins, Processes, Treatment and Outcome. New York: Oxford University Press, pp. 17-30.
Goldberg, G., and K. K. Bloom. (1990). The alien hand sign. American Journal of Physical Medicine and Rehabilitation 69(5):228-238.
Houk, J. C. (1995). Information processing in modular circuits linking basal ganglia and cerebral cortex. In J. C. Houk, J. L. Davies, and D. G. Beiser, Eds., Model of Information Processing in the Basal Ganglia. Cambridge, MA: Bradford, pp. 3-10.
Kosslyn, S. M. (1994). Image and Brain. Cambridge, MA: MIT Press.
Norman, D. A., and T. Shallice. (1986). Attention to action: willed and automatic control of behavior. In R. J. Davidson, G. E. Schwartz, and D. Shapiro, Eds., Consciousness and Self Regulation. New York: Plenum, pp. 1-17.
Petersen, S. E., P. T. Fox, M. I. Posner, M. Mintun, and M. E. Raichle. (1989). Positron emission tomographic studies of the processing of single words. Journal of Cognitive Neuroscience 1:153-170.
Posner, M. I., and G. J. DiGirolamo. (1998). Conflict, target detection and cognitive control. In R. Parasuraman, Ed., The Attentive Brain. Cambridge, MA: MIT Press.
Posner, M. I., and M. E. Raichle. (1994). Images of Mind. New York: Scientific American Library.
Rayner, K., and S. C. Sereno. (1994). Eye movements in reading: psycholinguistic studies. In M. A. Gernsbacher, Ed., Handbook of Psycholinguistics. New York: Academic Press, pp. 57-81.
Smith, E. E., and J. Jonides. (1995). Working memory in humans: neuropsychological evidence. In M. S. Gazzaniga, Ed., The Cognitive Neurosciences. Cambridge, MA: MIT Press, pp. 1009-1020.
Snyder, A. Z., Y. Abdullaev, M. I. Posner, and M. E. Raichle. (1995). Scalp electrical potentials reflect regional cerebral blood flow responses during processing of written words. Proceedings of the National Academy of Sciences 92:1689-1693.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. Journal of Experimental Psychology 18:643-662.
Toga, A. W., and J. C. Mazziotta, Eds. (1996). Brain Mapping: The Methods. New York: Academic Press.
Bisiach, E. (1992). Understanding consciousness: clues from unilateral neglect and related disorders. In A. D. Milner and M. D. Rugg, Eds., The Neuropsychology of Consciousness. London: Academic Press, pp. 113-139.
Burgess, P. W., and T. Shallice. (1996). Response suppression, initiation and strategy use following frontal lobe lesions. Neuro-psychologia 34:263-273.
Chelazzi, L., E. K. Miller, J. Duncan, and R. Desimone. (1993). A neural basis for visual search in inferior temporal cortex. Nature 363:345-347.
D"Esposito, M., J. A. Detre, D. C. Alsop, R. K. Shin, S. Atlas, and M. Grossman. (1995). The neural basis of the central executive system of working memory. Nature 378:279-281.
Démonet, J. F., R. Wise, and R. S. J. Frackowiak. (1993). Language functions explored in normal subjects by positron emission tomography: a critical review. Human Brain Mapping 1:39-47.
Graves, R. E., and B. S. Jones. (1992). Conscious visual perceptual awareness vs. non-conscious visual spatial localisation examined with normal subjects using possible analogues of blindsight and neglect. Cognitive Neuropsychology 9(6):487-508.
Jonides, J. P. (1981). Voluntary versus automatic control over the mind"s eye. In J. Long and A. Baddeley, Eds., Attention and Performance IX. Hillsdale, NJ: Erlbaum, pp. 187-204.
LaBerge, D. (1995). Attentional Processing: The Brain"s Art of Mindfulness. Cambridge, MA: Harvard University Press.
Pardo, J. V., P. T. Fox, and M. E. Raichle. (1991). Localization of a human system for sustained attention by positron emission tomography. Nature 349(6304):61-64.
Pardo, J. V., P. J. Pardo, K. W. Janer, and M. E. Raichle. (1990). The anterior cingulate cortex mediates processing selection in the Stroop attentional conflict paradigm. Proceedings of National Academy of Science 87:256-259.
Posner, M. I., G. J. DiGirolamo, and D. Fernandez-Duque. (1997). Brain mechanisms of cognitive skills. Consciousness and Cognition 6:267-290.
Rafal, R. D. (1994). Neglect. Current Opinion in Neurobiology 4:231-236.
Stuss, D. T., T. Shallice, M. P. Alexander, and T. W. Picton. (1995). A multidisciplinary approach to anterior attention functions. In J. Grafman, K. J. Holyoak, and F. Boller, Eds., Structure and Functions of the Human Prefrontal Cortex. New York: New York Academy of Sciences.
Umiltà, C. (1988). Orienting of attention. In F. Boller and J. Grafman, Eds., Handbook of Neuropsychology. Amsterdam: Elsevier, pp. 175-192
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology