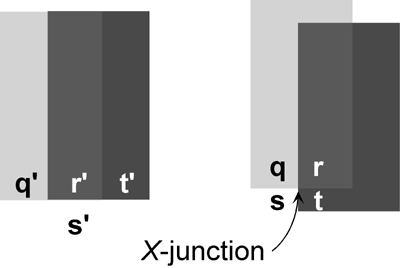
Figure 1
The light projecting to a given point in the RETINA possesses but a single value of color and intensity. When transparency is perceived, however, this light is interpreted as being reflected off of two (or sometimes more) surfaces lying in different depth planes. Perceptual transparency is a type of SURFACE PERCEPTION and illustrates the visual system's remarkable ability to reconstruct the three spatial dimensions of the environment given a stimulus (i.e., the retinal images) with only two. There are an infinite number of possible environmental causes of any particular pattern of retinal stimulation. The perception of transparency relies, as does visual perception in general, upon context to determine the most likely interpretation. For example, whereas region r" in figure 1 (left) is usually interpreted in terms of the color of a single surface, region r(right), an identical shade of gray, is seen to arise from light reflected off of two surfaces. This difference in perceptual interpretation is due to the presence of a contextual cue known as an X-junction.
X-junctions are the single most important monocular cue for transparency. They are defined by the presence of four contiguous regions (q,r,s,t; see figure 1) of an image with a characteristic spatial arrangement. Psychophysical studies have shown that the intensity relationships between these four regions must lie within certain bounds for perceptual transparency to occur. When X-junctions elicit a perception of transparency, two regions (q and s in figure 1) are seen as differently colored parts of the unoccluded background and the other two regions (r and t in figure 1) appear to be viewed through a foreground transparent surface (the darker rectangle). Perceptual psychologists have developed several simple physical models to account for the perception of transparency (e.g., Beck et al. 1984; Matelli 1985). Though differing slightly in their details, the optical properties of transmittance and reflectance are generally invoked in these models.
Transmittance refers to the multiplicative attenuation of background intensity. One way to think of transmittance is to imagine that transparent surfaces are generally opaque but have holes (like a fine wire mesh) too small to resolve (Kersten 1991; Richards and Witkin 1979; Stoner and Albright 1996). Transmittance is then the proportion of the surface with holes. Reflectance, on the other hand, refers to the fraction of incident light reflected off of a surface. If the surface is a foreground transparent surface, this light adds to that reflected off of the background surface. X-junctions that elicit a sense of transparency are usually those in which the four sub-regions possess intensities consistent with physically realizable values of transmittance and reflectance, giving credence to the idea that the visual system possesses a tacit model of the physics of transparency. Given the relative rarity of transparent surfaces in natural scenes, however, it might seem puzzling that the visual system would devote neural machinery to its detection. An alternative possibility is that perceptual transparency depends upon mechanisms that typically process the more common visual phenomena of shadows and opaque occlusion.Shadows and opaque occlusion are ubiquitous in natural scenes and, moreover, can be thought of as defining limiting cases of perceptual transparency.
Objects that lie between a light source and another object cast shadows such that the intensities of the shadowed regions appear multiplicatively diminished relative to the unshadowed regions. Shadows yield X-junctions that can be mimicked by a transparent surface possessing a transmittance less than one and having zero reflectance. Because the changes in intensities are purely multiplicative, the contrast (defined here as the intensity of one region divided by the intensity of another) between two adjacent regions in shadow should be the same as the contrast between spatially contiguous regions that lie out of shadow. Thus, if the X-junction in figure 1 were due to shadow-like transparency, the contrast between "unshadowed" regions q and s would be equal to that between their corresponding "shadowed" regions r and t.
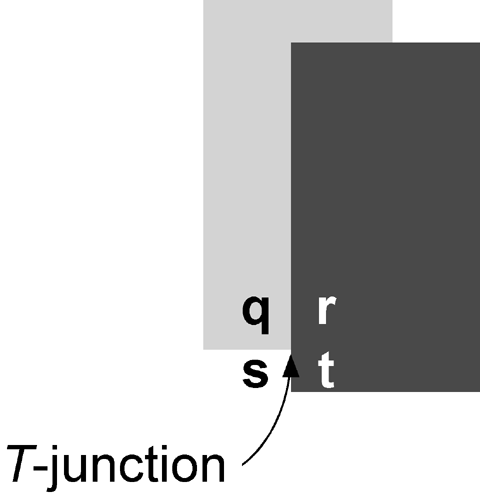
Opaque occlusion, on the other hand, results in total contrast attenuation -- regions r and t of figure 2 have identical intensities and hence no contrast exists between them. Opaque occlusion is associated with the presence of a contextual cue known as a T-junction. T-junctions are defined by the presence of three adjacent regions of differing intensity. In figure 2, these regions are q, s, and r/t (which form a single continuous region). The contrast reduction associated with opaque occlusion provides a very important cue for the depth ordering between surfaces. Indeed, the strength of the depth-ordering cue provided by transparent occlusion is directly proportional to the degree of contrast reduction with opaque occlusion providing the most potent cue. X- junctions associated with multiplicative transparency and T-junctions define two ends of a continuum within which perceptual transparency is most likely. Given this tight association between the stimulus conditions defining shadows, transparency, and opaque occlusion, it is tempting to speculate that transparency is detected by neural mechanisms that evolved to process shadows and opaque occlusion rather than transparency.
The neural mechanisms underlying the perception of transparency are only beginning to be understood. There are two important facets of this process. One is the detection of X-junctions suggestive of transparent overlap. As of this writing, no one has looked explicitly for neuronal responses to X-junctions. Baumann, van der Zwan, and Peterhans (1997) have, however, discovered neurons in area V2 of the macaque monkey that appear to encode the depth-ordering information implied by T-junctions. Whether, as the observations made above suggest, these neurons also respond selectively to X-junctions awaits an answer.
A second important facet of the neural processing of transparency is the existence of a multivalued representation of a particular surface property. For static displays there are at least two attributes that have multivalued perceptual representations -- surface reflectance and depth. The neural correlates of these multivalued representations have yet to be identified. For dynamic stimuli, more than one motion may be seen to project upon the same point in the retinal images. It has been shown that the perception of "motion transparency" interacts with the perception of transparency of these other attributes. Thus, when overlapping gratings moving in different directions are superimposed ("plaid patterns") in a manner that produces X-junctions which elicit a percept of transparency in static images, a perception of motion transparency (also known as "motion noncoherency") results: the two gratings are seen to move independently (Stoner, Albright, and Ramachandran 1990). Conversely, when plaid patterns have X-junctions inconsistent with transparent overlap, motion transparency is unlikely to occur. Neural correlates for motion transparency have been located (Stoner and Albright 1992). When stimulated with perceptually transparent moving plaid patterns, each of the two gratings activates a separate population of directionally tuned neurons in area MT. Conversely, when stimulated with nontransparent stimuli, a single population of MT neurons, sensitive to the composite motion of the plaid pattern, is activated. A plausible neural substrate for this type of transparency, at least, therefore appears to have been identified.
Beck, J., K. Prazdny, and R. Ivry. (1984). The perception of transparency with achromatic colors. Perception and Psychophysics 35:407-422.
Baumann R., R. van der Zwan, and E. Peterhans. (1997). Figure-ground segregation at contours: A neural mechanism in the visual cortex of the alert monkey. Eur. J. Neurosci. 9(6):1290-1303.
Kersten, D. (1991). Transparency and the cooperative computation of scene attributes. In M. Landy and A. Movshon, Eds., Computational Models of Visual Processing. Cambridge, MA: MIT Press, pp. 209-228.
Matelli, F. (1985). Stimulation and perception of transparency. Psychological Research 7:185-202.
Richards, W., and A. P. Witkin. (1979). Efficient Computations and Representations of Visible Surfaces. Tech. Rep. AFOSR-79-0020. Cambridge, MA: MIT.
Stoner, G. R., and T. D. Albright. (1992). Neural correlates of perceptual motion coherence. Nature 358:412-414.
Stoner, G. R., and T. D. Albright. (1996). The interpretation of visual motion: Evidence for surface segmentation mechanisms. Vision Research 36:1291-1310.
Stoner, G. R., T. D. Albright, and V. S. Ramachandran. (1990). Transparency and coherence in human motion perception. Nature 344:153-155.
Anderson, B. L. (1997). A theory of illusory lightness and transparency in monocular and binocular images: The role of contour junctions. Perception 26(4):419-453.
Gilchrist, A. L. (1994). Lightness, Brightness, and Transparency. Hillsdale, NJ: Erlbaum.
Kersten, D., H. H. Balthoff, B. L. Schwartz, and K. J. Kurtz. (1992). Interaction between transparency and structure from motion. Neural Computation 4:573-589.
Masin, S. C. (1997). The luminance conditions of transparency. Perception 26(1):39-50.
Nakayama, K., S. Shimojo, and V. S. Ramachandran. (1990). Transparency: Relation to depth, subjective contours, luminance, and neon color spreading. Perception 19(4):497-513.
Snowden, R. J., S. Treue, R. G. Erickson, and R. A. Andersen. (1991). The response of area MT and V1 neurons to transparent motion. J. Neurosci. 11(9):2768-2785.
Stoner, G. R., and T. D. Albright. (1993). Image segmentation cues in motion processing: Implications for modularity in vision. Journal of Cognitive Neuroscience 5:129-149.
Trueswell, J. C., and M. M. Hayhoe. (1993). Surface segmentation mechanisms and motion perception. Vision Res. 33(3):313-328.
Watanabe T., and P. Cavanagh. (1993). Surface decomposition accompanying the perception of transparency. Spatial Vision 7(2):95-111 .
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology