Visual scenes typically contain more objects than can ever be recognized or remembered in a single glance. Some kind of sequential selection of objects for detailed processing is essential if we are to cope with this wealth of information. Built into the earliest levels of vision is a powerful means of accomplishing the selection, namely, the heterogeneous RETINA. Fine grain visual resolution is possible only within the central retinal region known as the fovea, whose diameter is approximately 2 degrees of visual angle (about the size of eight letters on a typical page of text). Eye movements are important because they bring selected images to the fovea, and also keep them there for as long as needed to recognize the object.
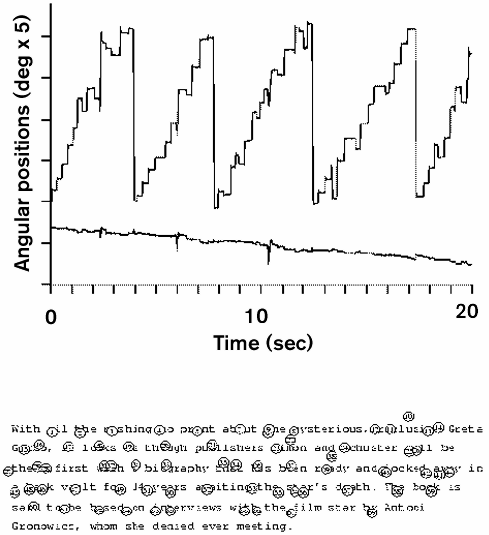
Eye movements fall into two broad classes. Saccadic eye movements (saccades) are rapid jumps of the eye used to shift gaze to any chosen object. In READING, for example, saccades typically occur about three times each second and are generally made to look from one word to the next (see figure 1). Smooth eye movements keep the line of sight on the selected object during the intervals between saccades, compensating for motion on the retina that might be caused either by motion of the object or by motion of the head or body. Intervals between saccades can be as long as several seconds during steady fixation of stationary or moving objects. Saccades can be made in any chosen direction, even in total darkness, whereas directed smooth eye movements cannot be initiated or maintained without some kind of motion signal.
Figure 1 Sequence of saccadic eye movements during reading. The graph on top shows horizontal (top trace) and vertical (bottom trace) eye movements over time. The abrupt changes in eye position are the saccades. The figure shows the sequences of rightward saccades (upward deflections in the trace) made to read a line of text, followed by large leftward resetting saccades made to the beginning of each successive line of text. The locations of the saccadic endpoints are shown in numbered sequence, superimposed on the text, at the bottom of the figure. The figure was made by J. Epelboim from recordings made with R. Steinman's Revolving Magnetic Field Sensor Coil monitor at the University of Maryland (see Epelboim et al. 1995, for a description of the instrument).
There are two natural links between eye movements and visual attention. One is the role played by ATTENTION in OCULOMOTOR CONTROL. The other is the way in which eye movements provide overt indicators of the locus of attention during performance of complex cognitive tasks, such as reading or visual search.
Consider first the role of attention in programming smooth eye movements. When we walk forward in an otherwise stationary scene, trying to keep our gaze fixed on our goal ahead, the flow of image motion generated on the retina by our own forward motion creates a large array of motion signals that could potentially drag the line of sight away from its intended goal (a problem described originally by Ernst Mach in 1906). Laboratory simulations of this common situation show that smooth eye movements can maintain a stable line of sight on a small, attended, stationary target superimposed on a large, vivid moving background. Similarly, smooth eye movements can accurately track a target moving across a stationary background. With more complex stimuli (letters, for example), perceptual identification of tracked targets is better than identification of untracked backgrounds (a result that holds after any differences in identification due to different retinal velocities of target and background are taken into account). The greater perceptibility of the target compared to the background implies that the same attentional mechanism serves both perception and eye movements (Khurana and Kowler 1987).
Attention contributes to the control of saccades in an analogous way, namely, attention is allocated to the chosen target shortly before the saccade is made to look at it (Hoffman and Subramaniam 1995; Kowler et al. 1995). Some attention can be transferred to nontargets, with no harmful effect on the latency or accuracy of the eye movements, showing that the attentional demands of eye movements are modest (Kowler et al. 1995; Khurana and Kowler 1987).
On the whole, the arrangement is very efficient. By allowing oculomotor and perceptual systems to share a common attentional filter, the eye will be directed to the object we are most interested in without the need for a separate selective attentional decision. At the same time, the modest attentional requirements of effective oculomotor control mean that it is very likely that we can look wherever we choose with little danger of the eye's being drawn to background objects, regardless of how large, bright, or vivid they may be. Modest attentional requirements also imply that there will be ample cognitive resources left over for identification and recognition; all our efforts need not be devoted to targeting eye movements.
The close link between attention and eye movements is supported by neurophysiology. Cortical centers containing neurons that are active before eye movements also contain neurons (sometimes the same ones) that are active before shifts of attention while the eye is stationary (Colby and Duhamel 1996; Andersen and Gnadt 1989). Some have gone so far as to consider whether shifting attention to an eccentric location while the eye remains stationary is equivalent to planning a saccadic eye movement (Kustov and Robinson 1997; Rizzolatti et al. 1987; Klein 1980).
Attention is involved in the programming of eye movements, and at the same time observations of eye movements provide a record of where someone chooses to attend during performance of complex cognitive tasks. Yarbus's (1967) well-known recordings of eye movements made while inspecting various paintings show systematic preferences to repeatedly look at those elements that would seem to be most relevant to evaluating the content of the picture. Despite the detailed record of preferences that eye movements provide, it has nevertheless proven to be surprisingly difficult to develop valid models of underlying cognitive processing based on eye movements alone (Viviani 1990). More recent work has taken a different tack by using highly constrained and novel tasks. Sequences of fixations have been used to study the modularity of syntactic processing during reading (Tanenhaus et al. 1995), the role of WORKING MEMORY during visual problem-solving tasks (Ballard, Hayhoe, and Pelz 1995; Epelboim and Suppes 1996), the coordination of eye and arm movements (Epelboim et al. 1995), and the size of the effective processing region during reading or search (McConkie and Rayner 1975; Motter and Belky 1998; O'Regan 1990).
This article has emphasized the importance of eye movements for selecting a subset of the available information for detailed processing. The price paid for having this valuable tool is that the visual system must cope with the continual shifts of the retinal image that eye movements will produce. Remarkably, despite the retinal perturbations, the visual scene appears stable and unimpaired. Evidence from studies in which subjects look at or point to targets presented briefly during saccades suggests that stored representations of oculomotor commands ("efferent copies") are used to take the effect of eye movements into account and create a representation of target location with respect to the head or body (Hansen and Skavenski 1977). Other evidence suggests that shifts of the retinal image are effectively ignored. According to these views, visual analysis begins anew each time the line of sight arrives at a target, with attended visual information converted rapidly to a high-level semantic code that can be remembered across sequences of saccades (e.g., O'Regan 1992).
The advantages to visual and cognitive systems of having a fovea are evidently so profound that it has been worth the cost of developing both the capacity for accurate control of eye movements and a tolerance for the retinal perturbations that eye movements produce. Visual attention is crucial for accomplishing both.
Andersen, R. A., and J. W. Gnadt. (1989). Posterior parietal cortex. In R. H. Wurtz and M. E. Goldberg, Eds., Reviews of Oculomotor Research, vol. 3: The neurobiology of saccadic eye movements. Amsterdam: Elsevier, pp. 315-336.
Ballard, D. H., M. M. Hayhoe, and J. B. Pelz. (1995). Memory representation in natural tasks. Journal of Cognitive Neuroscience 7:66-80.
Colby, C. L., and J.-R. Duhamel. (1996). Spatial representations for action in parietal cortex. Cognitive Brain Research 5:105-115.
Epelboim, J., R. M. Steinman, E. Kowler, M. Edwards, Z. Pizlo, C. J. Erkelens, and H. Collewijn. (1995). The function of visual search and memory in sequential looking tasks. Vision Research 35:3401-3422.
Epelboim, J., and P. Suppes. (1996). Window on the mind? What eye movements reveal about geometrical reasoning. Proceedings of the Cognitive Science Society 18: 59.
Hansen, R. H., and A. A. Skavenski. (1977). Accuracy of eye position information for motor control. Vision Research 17:919-926.
Hoffman, J., and B. Subramaniam. (1995). Saccadic eye movements and visual selective attention. Perception and Psychophysics 57:7787-7795.
Khurana, B., and E. Kowler. (1987). Shared attentional control of smooth eye movements and perception. Vision Research 27:1603-1618.
Klein, R. (1980). Does oculomotor readiness mediate cognitive control of visual attention? In R. Nickerson, Ed., Attention and Performance III. Hillsdale, NJ: Erlbaum, pp. 259-276.
Kowler, E. (1990). The role of visual and cognitive processes in the control of eye movement. In E. Kowler, Ed., Reviews of Oculomotor Research, vol. 4: Eye Movements and Their Role in Visual and Cognitive Processes. Amsterdam: Elsevier, pp. 1-70.
Kowler, E., E. Anderson, B. Dosher and E. Blaser. (1995). The role of attention in the programming of saccades. Vision Research 1897-1916.
Kustov, A. A., and D. L. Robinson. (1997). Shared neural control of attentional shifts and eye movements. Nature 384:74-77.
Mach, E. (1906/1959). Analysis of Sensation. New York: Dover.
McConkie, G. W., and K. Rayner. (1975). The span of the effective stimulus during a fixation in reading. Perception and Psychophysics 17:578-586.
Motter, B. C., and E. J. Belky. (1998). The zone of focal attention during active visual search. Vision Research 38:1007-1022.
O'Regan, J. K. (1990). Eye movements and reading. In E. Kowler, Ed., Reviews of Oculomotor Research, vol. 4: Eye Movements and Their Role in Visual and Cognitive Processes. Amsterdam: Elsevier, pp. 395-453.
O'Regan, J. K. (1992). Solving the "real" mysteries of visual perception: the world as an outside memory. Canadian Journal of Psychology 46:461-488.
Rizzolatti, G., L. Riggio, I. Dascola, and C. Umita. (1987). Reorienting attention across the horizontal and vertical meridians: evidence in favor of a premotor theory of attention. Neuropsychologia 25:31-40.
Tanenhaus, M. K., M. J. Spivey-Knowlton, K. M. Eberhard, and J. C. Sedivy. (1995). Integration of visual and linguistic information in spoken language comprehension. Science 268:1632-1634.
Viviani, P. (1990). Eye movements in visual search: cognitive, perceptual and motor control aspects. In E. Kowler, Ed., Reviews of Oculomotor Research, vol. 4: Eye Movements and Their Role in Visual and Cognitive Processes. Amsterdam: Elsevier, pp. 353-393.
Yarbus, A. L. (1967). Eye Movements and Vision. New York: Plenum Press.
Hoffman, J. E. (1997). Visual attention and eye movements. In H. Pashler, Ed., Attention. London: University College London Press.
Kowler, E. (1995). Eye movement. In S. Kosslyn, Ed., Invitation to Cognitive Science, vol. 2. Cambridge, MA: MIT Press, pp. 215-265.
Rayner, K, Ed. (1992). Eye Movements and Visual Cognition: Scene Perception and Reading. New York: Springer.
Steinman, R. M., and J. Z. Levinson. (1990). The role of eye movement in the detection of contrast and spatial detail. In E. Kowler, Ed., Reviews of Oculomotor Research, vol. 4: Eye Movements and Their Role in Visual and Cognitive Processes. Amsterdam: Elsevier, pp. 115-212
Suppes, P. (1990). Eye movement models for arithmetic and reading performance. In E. Kowler, Ed., Reviews of Oculomotor Research, vol. 4: Eye Movements and Their Role in Visual and Cognitive Processes. Amsterdam: Elsevier, pp. 455-477.
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology