This article concerns shape perception, so it is natural to consider what is meant by "shape." Most readers undoubtedly have an intuitive feel for the meaning of the term, sensing its relatedness to such concepts as form and structure. Nevertheless, a precise definition of shape has proved elusive. Instead, experimenters have adopted the working definition that shape is an aspect of a stimulus that remains invariant despite changes in size, position, and orientation. For example, 2-D visual stimuli have the same shape if there exists a transformation of spatial scale (e.g., magnification) or a rotation in the picture plane that renders them identical. Similarly, 3-D objects have the same shape if their volumes can be equated by size changes or a combination of rotations about three spatial axes.
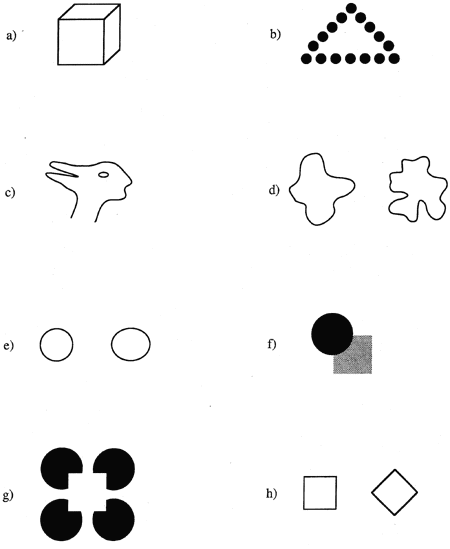
Although our world is filled with objects and patterns, most studies of shape perception have used only 2-D stimuli. This emphasis reflects the belief that perception of 3-D form depends on the shape of 2-D regions in the retinal image. The fact that line drawings can evoke vivid percepts of 3-D form (see figure 1a) supports the idea that 2-D and 3-D shape perception are related, but how they are related is still debated (cf. Attneave 1954; GIBSON 1950; Hochberg 1964; Koffka 1935; MARR 1982). Furthermore, variables that affect LIGHTNESS PERCEPTION (e.g., shading) and DEPTH PERCEPTION (e.g., binocular disparity and relative motion) also affect form perception. How these variables interact with 2-D shape to yield a 3-D percept is not well understood.
Figure 1 (a) Although this image is two-dimensional, observers perceive a three-dimensional cube.
(b). Shape perception can occur after perceptual grouping. Observers can perceive both the global (triangle) and the local (circle) shapes.
(c). Perceived shape is influenced by top-down processes. When one stimulus has multiple interpretations, perceived shape changes even with no stimulus change. This figure can be seen as a duck (facing left) or as a rabbit (facing right). When the percept changes, so does the interpretation of specific parts of the stimulus (e.g., what was a beak becomes ears).
(d). We perceive shapes of unfamiliar figures that have no a priori meaning. Observers can discriminate these two shapes, although we have no memory association for either.
(e). Quite small shape differences can be perceived. The length of this oval is just 20% longer than the diameter of the circle. Most observers can discriminate even more subtle differences in shape.
(f). Shape perception proceeds even in the absence of complete stimulus information. This figure is consistent with both a circle partially occluding a square (in two depth planes), and a circle next to a notched square (in one or more depth planes). Most people perceive the former -- filling in missing shape information.
(g). We can perceive the shapes of completely invisible objects. In this example of an illusory figure, observers see a white square on top of four black circles. Although the edges of the square appear highly salient, they do not physically exist.
(h). Mach's original demonstration that phenomenological shape can vary with orientation. The two stimuli have the same physical shape, but observers perceive the left shape as a square and the right as a diamond (if the reader tilts his or her head 45 degrees clockwise, the percepts reverse).
In most viewing conditions, a pattern's shape is closely related to the spatial arrangement of its contours. Thus, shape perception depends in part on feature extraction processes that encode edges and elementary features in the retinal image, and on processes that group elements into higher-order units (see GESTALT PERCEPTION). In the latter case, we can perceive both the shape of the individual elements, as well as the global shape of the grouped elements (figure 1b). Despite the close connection between contours and shape, simultaneous presentation of contours is neither necessary nor sufficient for shape perception. It is not necessary because shape is perceived even when only isolated parts of contour are presented. For example, in anorthoscopic stimuli a visual pattern is moved behind a stationary slit in an opaque screen: although only a small dot or line is visible at any one time, observers often perceive the shape of the hidden pattern. Simultaneous presentation of contours is not sufficient for shape perception because observers do not necessarily attend to everything in the visual field. Although some attributes of an unattended object, such as its presence, location, and color, can be perceived, ATTENTION appears to be required for shape perception (Rock and Gutman 1981; Rock et al. 1992). Other top-down processes also affect shape perception, as is shown by ambiguous figures like the duck/rabbit pattern in figure 1c: The contours remain constant, but perceived shape depends on the interpretation of the figure. In some cases, the allocation of attention to particular stimulus features can bias the percept of ambiguous figures (Peterson and Gibson 1991).
Although clearly related to HIGH-LEVEL VISION phenomena such as OBJECT RECOGNITION, shape perception is better classified as an aspect of MID-LEVEL VISION because the perception of shape does not require recognition: One can perceive the shapes of novel stimuli that have no a priori meaning (figure 1d), and an observer can recognize objects based on surface properties like TEXTURE or color (Humphrey et al. 1994; see also SURFACE PERCEPTION). Nevertheless, shape provides important clues about an object's identity, as well as information that is critical to manipulating objects and determining their functional properties or AFFORDANCES (e.g., Can I stand on this object?). Thus, it is not surprising that visual mechanisms encode shape rapidly and accurately. For example, the response time needed to name line drawings of familiar objects, or to determine if two novel polygons have the same shape, is approximately 1 sec (Larsen 1985). Although these response times are short, they probably overestimate the time needed to perceive shape. For instance, observers can identify letters and line drawings of familiar objects presented for only 50 msec (Biederman and Ju 1988; Jolicoeur and Landau 1984). Other studies have shown that observers are exquisitely sensitive to shape differences that are much more subtle than those shown in figure 1e, even when the stimuli being compared differ substantially in size (De Valois et al. 1990; Regan and Hamstra 1992).
Further evidence of the versatility of shape perception comes from the fact that a stimulus need not be completely visible for observers to perceive shape accurately. In the everyday world, objects frequently occlude parts of neighboring objects, yet our visual systems are able to complete missing shape information (figure 1f). Although shape completion appears effortless and immediate, it is not instantaneous, and the way in which shapes are completed depends on factors such as stimulus regularity and symmetry (Sekuler 1994; Sekuler and Palmer 1992). A phenomenon related to visual completion is that of illusory contours, in which the percept of a shape is induced by other visible shapes (figure 1g; Kanizsa 1975).
At the beginning of this article we defined shape as that aspect of a stimulus that remained invariant despite changes in size and orientation. Thus, one might expect that perceived shape would not depend on stimulus orientation or size. This suggestion seems intuitively obvious, and it is consistent with our experience in everyday tasks in which objects are recognized (in part on the basis of shape) despite substantial changes in retinal size and orientation. Nevertheless, there is ample evidence that our perception of shape is not invariant across spatial scales and orientations. For example, it is more difficult to recognize line drawings of familiar objects and abstract patterns presented in unfamiliar sizes or orientations (Edelman and Bülthoff 1992; Jolicoeur 1987; Rock, Schreiber, and Ro 1994; Tarr and Pinker 1989). Furthermore, it takes significantly more time to determine if two patterns have the same shape when they differ in size or orientation (Larsen 1985; Sekuler and Nash 1972). Finally, there are several demonstrations that phenomenological shape varies significantly with orientation. Mach's classic demonstration of the effect of orientation on the perceived shape of a square is shown in figure 1h. In Mach's words, "Two figures may be geometrically congruent, but physiologically quite different, . . . [these figures] could never be recognized as the same without mechanical and intellectual operations" (Mach 1959: 106; see also MENTAL ROTATION). These effects of size and orientation on shape perception have important implications for theories of object recognition. Specifically, they raise the possibility that an object is recognized by comparing it to multiple, viewpoint-dependent representations in long-term memory, rather than to a single, viewpoint-independent representation.
Relatively little is known about the physiological mechanisms underlying shape perception, but researchers have located neurons in the inferotemporal cortex (IT) that respond better to some shapes than to others, even when these shapes are not identified with specific previously learned objects. Whereas the primary VISUAL CORTEX initially codes basic features such as orientation, size, and color, area IT codes much more complex features such as particular shapes, or combinations of shapes and colors (Tanaka 1993). Neurons in area IT also have similar shape selectivity regardless of the cue that defines the shape (e.g., luminance-, texture-, or motion-defined shapes; Sary et al. 1995), the size and position of the shape (Ito et al. 1995; Logothetis, Pauls, and Poggio 1995), or the presence or absence of partially occluding contours (Kovacs, Vogels, and Orban 1995). These recent discoveries suggest that area IT plays a significant role in shape constancy and recognition, but other cortical areas may be involved in the use of shape to guide the manipulation of objects (Goodale and Milner 1992; see also VISUAL PROCESSING STREAMS). Additional physiological and psychophysical research in the next few years undoubtedly will increase our understanding of the physiological processes underlying shape perception.
Attneave, F. (1954). Some informational aspects of visual perception. Psychological Review 61:183-193.
Biederman, I., and G. Ju. (1988). Surface versus edge-based determinants of visual recognition. Cognitive Psychology 20:38-64.
De Valois, K., V. Lakshminarayanan, R. Nygaard, S. Schlussel, and J. Sladky. (1990). Discrimination of relative spatial position. Vision Research 30:1649-1660.
Edelman, S., and H. Bülthoff. (1992). Orientation dependence in the recognition of familiar and novel views of three-dimensional objects. Vision Research 32:2385-2400.
Gibson, J. J. (1950). Perception of the Visual World. Boston, MA: Houghton-Mifflin.
Goodale, M., and A. Milner. (1992). Separate visual pathways for perception and action. Trends in Neuroscience 15:20-25.
Hochberg, J. E. (1964). Perception. Englewood Cliffs, NJ: Prentice-Hall.
Humphrey, G., M. Goodale, L. Jakobson, and P. Servos. (1994). The role of surface information in object recognition: Studies of a visual form agnosic and normal subjects. Perception 23:1457-1481.
Ito, M., H. Tamura, I. Fujita, and K. Tanaka. (1995). Size and position invariance of neuronal responses in monkey inferotemporal cortex. Journal of Neurophysiology 73:218-226.
Jolicoeur, P. (1987). A size-congruency effect in memory for visual shape. Memory and Cognition 15:531-543.
Jolicoeur, P., and M. Landau. (1984). Effects of orientation on the identification of simple visual patterns. Canadian Journal of Psychology 38:80-93.
Kanizsa, G. (1975). Contours without gradients or cognitive contours? Italian Journal of Psychology 1:93-112.
Koffka, K. (1935). Principles of Gestalt Psychology. New York: Harcourt, Brace, and World Inc.
Kovacs, G., R. Vogels, and G. Orban. (1995). Selectivity of macaque inferior temporal neurons for partially occluded shapes. Journal of Neuroscience 15:1984-1997.
Larsen, A. (1985). Pattern matching: Effects of size ratio, angular difference in orientation and familiarity. Perception and Psychophysics 38:63-68.
Logothetis, N., J. Pauls, and T. Poggio. (1995). Shape representation in the inferior temporal cortex of monkeys. Current Biology 5:552-563.
Mach, E. (1959). The Analysis of Sensations. Translated by C. M. Williams. New York: Dover Publishers.
Marr, D. (1982). Vision. San Francisco, CA: W. H. Freeman.
Peterson, M. A., and B. S. Gibson. (1991). Directing spatial attention within an object: Altering the functional equivalence of shape descriptions. Journal of Experimental Psychology: Human Perception and Performance 17:170-182.
Regan, D., and D. Hamstra. (1992). Shape discrimination and the judgement of perfect symmetry: Dissociation of shape from size. Vision Research 32:1845-1864.
Rock, I., and D. Gutman. (1981). The effect of inattention on form perception. Journal of Experimental Psychology: Human Perception and Performance 7:275-285.
Rock, I., C. Linnett, P. Grant, and A. Mack. (1992). Perception without attention: Results of a new method. Cognitive Psychology 24:502-534.
Rock, I., C. Schreiber, and T. Ro. (1994). The dependence of two-dimensional shape perception on orientation. Perception 23:1389-1506.
Sary, G., R. Vogels, G. Kovacs, and G. Orban. (1995). Responses of monkey inferior temporal neurons to luminance-, motion- and texture-defined gratings. Journal of Neurophysiology 73:1341-1354.
Sekuler, A. (1994). Local and global minima in visual completion: Effects of symmetry and orientation. Perception 23:529-545.
Sekuler, A., and S. Palmer. (1992). Perception of partly occluded objects: A microgenetic analysis. Journal of Experimental Psychology: General 121:95-111.
Sekuler, R., and D. Nash. (1972). Speed of size scaling in human vision. Psychonomic Science 1972:93-94.
Tanaka, K. (1993). Neuronal mechanisms of object recognition. Science 262:685-688.
Tarr, M., and S. Pinker. (1989). Mental rotation and orientation-dependence in shape perception. Cognitive Psychology 21:233-282.
Biederman, I. (1987). Recognition-by-components: A theory of human image understanding. Psychological Review 94:115-147.
Cavanagh, P., and Y. Leclerc. (1989). Shape from shadows. Journal of Experimental Psychology: Human Perception and Performance 15:3-27.
Jolicoeur, P. (1992). Identification of disoriented objects: A dual-systems theory. In G. Humphreys, Ed., Understanding Vision: An Interdisciplinary Perspective. Cambridge, MA: Blackwell, pp. 180-198.
Marr, D., and H. Nishihara. (1978). Representation and recognition of the spatial organization of three-dimensional shapes. Proceedings of the Royal Society of London Series B Biological Sciences 200:269-294.
Rock, I. (1973). Orientation and Form. New York: Academic Press.
Rock, I. (1983). The Logic of Perception. Cambridge, MA: MIT Press.
Uttal, W. (1988). On Seeing Forms. Hillsdale, NJ: Erlbaum.
Zusne, L. (1970). Visual Perception of Form. New York: Academic Press.
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology