In primates visual processing is carried out in many different parts of the brain. Described here is the basic anatomy and physiology of the RETINA, the lateral geniculate nucleus of the THALAMUS, the striate cortex, higher cortical visual areas, the superior colliculus, and the accessory optic system (AOS).
The retina: Five major classes of neurons have been discerned in the primate retina: photoreceptors, horizontal cells, bipolar cells, amacrine cells, and ganglion cells. Several subclasses have been discerned within each of these cell types. The rod and cone photoreceptors subserve night and day vision. In humans and many primates three kinds of cones have been discerned that have maximum sensitivity for either short-(blue), medium-(green), or long-wavelength (red) light. All photoreceptors hyperpolarize to light, yield only graded potentials, and use the neurotransmitter glutamate. Bipolar cells form two major classes, the ON and OFF types, which by virtue of sign-inverting and sign-conserving synapses, respectively, depolarize and hyperpolarize to light. Several subclasses of ON and OFF bipolar cells have been identified. There are numerous classes of amacrine cells that use a variety of different neurotransmitters that include dopamine, acetylcholine, and serotonin. The ganglion cells are the output cells of the retina. Each ganglion cell is sensitive to a small area of the visual field, called the receptive field of the cell. Most commonly the shape of the receptive field is circular and consists of two regions, a central excitatory one which, when stimulated by light or dark spots (for the ON and OFF cells respectively), elicits a vigorous burst of action potentials in the cell, and a concentric inhibitory surround area produced largely by virtue of the horizontal cell network. As a result of this arrangement, responses arise predominantly when a local difference in luminance or chrominance is detected within the receptive field.
Two major classes of the primate ganglion cells are the midget and parasol cells. The midget ganglion cells, as their name implies, are quite small; in central retina their receptive field center is composed of but a single cone, rendering them thereby, in trichromatic primates, wavelength-selective. The parasol cells, by contrast, are much larger and receive input both in the center and surround of their receptive fields from several different cone types. As a result of convergent input from the receptors, the parasol cells are more sensitive to contrast but cannot provide information about differences in wavelength. Midget and parasol cells come in two subvarieties, the ON and the OFF. The ON cells are excited by light increment and the OFF cells are excited by light decrement. The midget system is believed to make a significant contribution to the processing of fine detail and color, and the parasol system to the processing of rapid motion and luminance changes. In central vision the midget cells outnumber the parasol cells ten to one. This ratio declines with increasing eccentricity; in the far periphery the two cells types are just about equally numerous. Another group of retinal ganglion cells receives convergent input from ON and OFF bipolar cells as a result of which they discharge to both light increment and light decrement. Such cells make extensive projections to the superior colliculus.
The optic nerve consists of the axons of the retinal ganglion cell. In animals with lateral eyes, such as fish and amphibians, most of the axons in the optic nerve of each eye cross over at the optic chiasm to the contralateral hemisphere. In higher mammals with forward-looking eyes, only the axons of ganglion cells in the nasal hemiretinae cross over at the chiasm; the temporal hemiretinae project ipsilaterally. This arrangement makes it possible for corresponding points in the retinae of the two eyes to reach the same sites in cortex to realize binocular vision.
The retinal ganglion cell axons terminate in several central structures that include the lateral geniculate nucleus of the thalamus, the superior colliculus (called the optic tectum in reptiles and amphibians), and the terminal nuclei.
The lateral geniculate nucleus of the thalamus: The two lateral geniculate nuclei, one on each side of the brain, are laminated thalamic structures. Each receives input from the nasal hemiretina of one eye and the temporal hemiretina of the other, thereby representing the contralateral visual hemifield. In primates there are six layers for central vision, three of which receive input from one eye and three from the other. The midget retinal ganglion cells project to the top four parvocellular layers, whereas the parasol cells project to the bottom two magnocellular layers. The number of layers reduces to four for peripheral representation, two of which are parvocellular and two magnocellular. The interlaminar layers contain small cells innervated by several classes of small retinal ganglion cells. The retinal projections are orderly, thereby creating a neat topographic arrangement in the structure. The receptive field properties of single cells are quite similar to those seen in retinal ganglion cells. Their responses, however, can be modulated by the extensive inputs the lateral geniculate nucleus receives directly or indirectly from the cortex.
The striate cortex: The striate cortex is an expanse of cortical tissue that in higher mammals comprises a large portion of the occipital lobe. Transverse sections of this tissue, when stained with cresyl violet, reveal a distinct striation in layer 4, called the stripe of Gennary; this stripe is not evident in other cortical regions and thereby provides an easy anatomical delineation of the area. The striate cortex, often called V1 (for visual 1), spans a thickness of approximately 2 mm of gray matter; six major laminae have been discerned, some of which have subsequently been divided into several sublaminae. In primates the striate cortex is the major recipient zone of the input from the dorsal lateral geniculate nucleus of the thalamus; fibers terminate most profusely, but not exclusively, in layer 4c, which has two subdivisions, 4cα and 4cβ. The inputs to 4cα and 4cβ arise, respectively, from the magnocellular and parvocellular divisions of the lateral geniculate nucleus.
The visual field is represented in an orderly manner along the surface extent of the striate cortex with more space allocated for central than for peripheral vision. Each cell in the striate cortex is sensitive to a relatively small region of the visual field, the receptive field area of the cell. By virtue of elaborate excitatory and inhibitory connections, the input from the lateral geniculate nucleus is reorganized to yield six major changes in receptive field properties: (1) selectivity for the orientation of edges, (2) selectivity for the direction of stimulus movement, (3) selectivity for the spatial frequency of repetitive stimuli such as textures, (4) selectivity for color along several directions in color space in addition to the red/green and blue/yellow axes seen in the retinal ganglion cells and in the lateral geniculate nucleus, (5) sensitivity for both light increment (ON response) and light decrement (OFF response) in many cells, and (6) selectivity for binocular activation that gives rise to stereoscopic depth perception. Orientation-specific cells in the striate cortex appear to form several distinct classes that include the so-called simple and complex cells. The receptive fields of simple cells subdivide into spatially separate regions, within each of which responses are produced for either light increment (ON response) or light decrement (OFF response) but not both. Complex cells, which commonly receive input from both eyes, do not exhibit such subdivision; they typically respond to both light increment and decrement throughout their receptive fields. Furthermore, their receptive fields are larger than those of simple cells.
In addition to the laminar organization of the striate cortex, columnar organization has also been revealed, most notably for orientation specificity. Within each column cells have the same orientation. The columns form stripes along the cortical surface with neighboring columns showing progressive shifts in orientation preference. This arrangement, coupled with columnar organization for ocular dominance, is modular. Each module, sometimes called a "hypercolumn," measures approximately 1 mm by 1 mm with a depth of 2 mm; it is conceived to be a self-contained processing unit for a given location of the visual field within which the basic attributes believed to be necessary for the analysis of the visual scene are contained. These include a full representation of neurons for the local analysis of orientation, direction, spatial frequency, color, and depth. The columnar organization seen in the striate cortex is believed to optimize encoding and visual analysis.
Higher cortical areas: In the primate posterior cortex more than thirty distinct visual areas that make more than 300 interconnections have been identified. These include areas V2, V3, V4, the middle temporal (MT), the medial superior temporal (MST), the ventral interparietal (VIP), and the inferior temporal (IT) areas. Most of these areas are also modular, as is area V1. Physiological studies have shown that the modules perform a great variety of analyses. The receptive fields of single cells become larger in size the further the area is removed from V1; this goes along with a gradual loss in the topographic order with which the visual field is laid out in each area. The occipitoparietal areas are believed to play an important role in the analysis of spatial relationships and motion, whereas the occipito-temporal areas are believed to play a role in object recognition and color vision. Lesion studies suggest that there is considerable redundancy in processing as selective removal of single extrastriate areas such as V3, V4, and MT does not produce highly specific or long-lasting deficits in the analysis of basic visual capacities such as motion, color, shape, and depth. However, lesions of temporal cortex produce severe deficits in object recognition and lesions of the parietal cortex, shorter-term deficits in spatial vision. At this stage of the research it is still not clearly understood why so many cortical visual areas have evolved in primates.
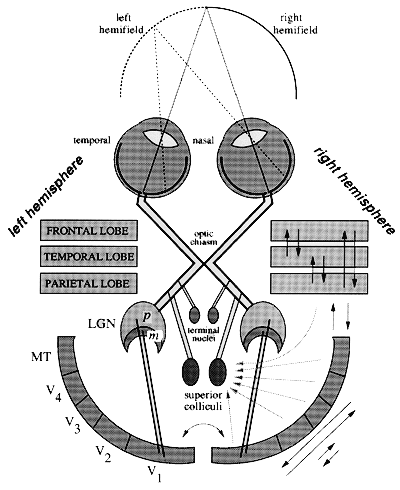
Figure 1 A schematic of the major structures and connections of the visual system as viewed from above. The temporal hemiretinae project ipsilaterally, whereas the projections from the nasal hemiretinae cross over at the optic chiasm to the contralateral hemisphere. As a consequence, corresponding retinal points in the two eyes project to the same hemisphere and to nearby regions within each target structure. Three retinal projections are depicted in the figure, which terminate in the lateral geniculate nuclei (LGN) of the thalamus, in the superior colliculi, and in the terminal nuclei. The magnocellular (m) and parvocellular (p) portions of the LGN project predominantly to primary visual cortex (V1). The extrastriate areas shown are V2, V3, V4, and MT. These areas are extensively interconnected, as indicated on the right. The temporal, parietal, and frontal lobes contain several additional visual areas that interconnect with each other as well as with the extrastriate areas. Most of these visual areas send projections to the superior colliculi as well as other subcortical visual centers.
The superior colliculus: The superior colliculus, located on the roof of the midbrain, is a laminated structure. In the upper layers the visual field is represented in an orderly fashion. Most cells have relatively small receptive fields and give both ON and OFF responses to stimuli; little selectivity has been reported for color and shape. In some species, like the cat for example, cells having directionally selective attributes are common. In the primates so far studied the percentage of such cells in the superior colliculus is low.
In the deeper layers of the primate superior colliculus single cells that respond in association with eye movements are common. Each cell discharges optimally when an eye movement of a certain direction and amplitude is executed; each region in the colliculus represents different directions and amplitudes of eye movements arranged in an orderly fashion. Electrical stimulation elicits saccadic eye movements with very low currents. The superior colliculus plays a central role in the generation of saccadic eye movements. However, several other structures, including regions in the occipital and parietal lobes, the frontal eye fields, and the medial eye fields in the frontal lobe also contribute to eye movement generation. Electrical stimulation in these areas also produces eye movements. Removal of the superior colliculus causes moderate deficits in eye movements, most notable being the inability to execute saccades to visual targets with very short latencies (the so-called express saccades). After colliculus removal, electrical stimulation of the occipital and parietal lobes no longer produces eye movements, although stimulation-elicited eye movements from the frontal areas remain unaffected. Removal of the frontal eye fields and the medial eye fields produces only minor deficits in eye movements. However, when both the frontal eye fields and the superior colliculi are removed, visually guided saccadic eye movements can no longer be generated. These findings suggest that there are two major systems for saccadic eye movement control: the posterior, which involves the occipital and parietal lobes that pass through the colliculus to reach the brain stem, and the anterior, which involves the frontal lobe that reaches the brain stem directly.
The accessory optic system: The AOS, extensively studied in the rabbit, arises from a special class of ganglion cells, the cells of Dogiel, that are directionally selective and respond best to slow rates of movement. They project to the terminal nuclei which in turn project to the dorsal cap of Kooy of the inferior olive. The climbing fibers from the olive project to the flocculo-nodular lobe of the cerebellum from where the brain stem oculomotor centers are reached through the vestibular nuclei.
The prime function of the accessory optic system appears to be to stabilize the retinal image by counterrotating the eyes when either the organism or the visual field is set in motion at slow velocities. At higher velocities retinal slip is prevented by the vestibular system which also sends its signals to the brain stem for eye movement generation via the vestibular nuclei.
Rockland, K. S., J. H. Kaas, and A. Peters, Eds. (1997). Extrastriate Cortex. Cerebral Cortex, vol. 12, Extrastriate Cortex. New York: Plenum Press.
Schiller, P. H. (1986). The central visual system. The 25th Jubilee Issue of Vision Research 26:1351-1386.
Spillman, L., and J. S. Werner, Eds. (1990). Visual Perception, The Neurophysiological Foundations. San Diego: Academic Press .
![]() Copyright © 1999 Massachusetts Institute of Technology
Copyright © 1999 Massachusetts Institute of Technology